
 Généralités
Généralités
Un plasmode est une masse cellulaire sans paroi.Les thalles unicellulaires sont trouvés, par exemple, chez les levures.
Les thalles pluricellulaires peuvent être à filaments non cloisonnés ou à filaments septés (on trouvera un ou plusieurs noyaux selon l’organisme).
La multiplication végétative peut se faire selon diverses modalités.
On peut trouver une multiplication par des organes spécialisés comme avec les sporocystes qui libèrent des spores. Les sporocystes sont portés par des sporocystophores (filament érigé). Les spores obtenus peuvent être de deux types :
- Des spores endogènes qui donneront des spores sèches ou des spores flagellées si elles sont dans une cellule spécialisée.
- Des spores exogènes qui sont aussi appelées des conidies. Dans ce cas, le filament porteur est un conidiophore.
- La planogamie.
Les gamétocystes (mâles ou femelles) donnent des gamètes flagellés qui sont émis à l’extérieur et qui vont fusionner. Cette fusion donne un planozygote qui se fixe. - L’oogamie.
Le gamétocyste mâle (ou spermatocyste) donne des gamètes flagellés (spermatozoïdes) qui vont aller féconder le gamète femelle (oosphère) dans le sporocyste femelle (oocyste). L’oeuf est contenu dans l’oocyste : c’est l’oospore. - L’oogamie siphonogame.
Le gamétocyste mâle ne donne pas de spermatozoïdes. Il doit venir s’accoler au gamétocyste femelle puis émettre des siphons copulateurs qui perforent la paroi de l’oocyste. - Cystogamie (ou zygogamie).
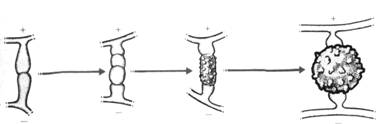
1 : progamétocystes + et – ;
2 : gamétocystes et suspenseurs ;
3 : fusion des gamétocystes ;
4 : zygospore.
Dans ce cas, on n’a besoin que de deux thalles compatibles, sans besoin de gamète. Il va y avoir émission d’un diverticule latéral (progamétocyste). Ensuite, apparaît une cloison latérale qui est le gamétocyste et le suspenseur.
Il va ensuite y avoir mélange des noyaux puis formation du zygote à paroi échinulée.
- La trichogamie.
Dans ce cas, le gamète mâle est une spermatie (non flagellé), émise par un filament.
L’organe femelle est un ascogone (cellule globuleuse surmontée d’un trichogyne). Il va y avoir fusion des parois puis injection du noyau mâle à l’intérieur de l’ascogone. Pendant le parcours du noyau mâle, ce dernier va se multiplier. Il y aura ensuite appariement sans fusion (dicaryon). On a alors des asques, aussi appelés, méiogamétocystes.
- La somatogamie.
Il va y avoir fusion de deux thalles compatibles et formations de dicaryons.
[table "5" not found /]
I Phylogénie des champignons.
C’est un règne à part comprenant environ 56 000 espèces connues (1 million d’espèces estimées). La classification est basée sur la morphologie, la biochimie et labiologie moléculaire.
A Les différents types de thalles.
Ils peuvent être macroscopiques souterrains (truffe) ou aériens (amanites). D’autres thalles sont microscopiques. Tous les champignons sont des eucaryotes, se nourrissant par absorption ; leur appareil végétatif est un thalle. Ils ne sont paschlorophylliens. On trouve les myxomycètes qui ont des mouvements amiboïdes car leur reproduction est réalisée par des sporocystes. On trouve aussi les oomycètes qui ont un
thalle filamenteux, une nutrition par absorption et une reproduction par formation de spores.
B Le rôle des champignons dans la nature.
Les champignons peuvent être : décomposeurs, symbiotiques, parasites d’animaux ou de végétaux.Ils sont une source d’alimentation pour l’Homme, soit de manière directe soit indirectement (fermentation, affinage).
L’identification se fait selon des critères morphologiques : on doit donc connaître plus ou moins la classification.
C Les différents types de thalles.
Le thalle est la partie végétative assurant l’absorption des nutriments et permettant le développement des organes reproducteurs. Parmi les différents thalles, on distingue :- Les plasmodes (myxomycètes)
- Les nématothalles : peuvent être érigés (allongés) ou coccoïdes.
- Les cladothalles : ils sont rares, comprennent les ascomycètes.
- Les thalles septés.
- Les thalles siphonnés : ont une cloison, pour délimiter un organe ou bine pour séparer une partie morte.
- Les agglomérats à hyphes : rigomorphes ou cordons (ressemblent à une racine) ; hyphes : cherchent les nutriments dans le sol, les parties anciennes s’agglomèrent pour former les hyphes.
- Les sclérotes : ce sont des amas d’hyphes dont des cellules de la périphérie se réunissent et les filaments centraux donnent la « moelle » riche en réserves nutritives. On les trouve chez les ascomycètes et chez les basidiomycètes. (Exemple : ergot du seigle).
- Les coussins conidifères : ce sont des agglomérats de conidiophores (Monilia laxa).
- Les acervules : ce sont les moins organisés. On les trouve sur les champignons de plantes. Les conidiophores s’accumulent sous l’épiderme du vaisseau (conidiophore).
- Les pycnides sont des structures en forme de bouteille avec un col qui forme un orifice.
- Les carpophores : ce sont des amas de filaments septés donnant un pied et un chapeau. Les hyphes sont serrés et forment un réseau lâche de filaments.
- Les stolons et rhizoïdes : ce sont des filaments qui poussent en ligne droite (stolon) puis qui se courbent. Il y a formation de rhizoïdes qui émettent une touffe de sporocystophores puis un nouveau stolon.
D Aspect cytologique du thalle.
Le noyau est petite taille, le nombre de chromosomes est restreint (entre 4 et 8) ; l’appareil de Golgi est atypique ; Présence de vacuoles dont l’importance augmente lors du vieillissement du thalle ; les réserves sont sous forme de lipides et de glycogène.E Composition de la paroi.
Elle est faite de 80% de polysaccharides, 10% de protéines et 10% de lipides. La chitine, chez les Eumycota est un polymère de N-actéyl glucose amine (β1–>4). Le chitosan est formé de glucose amine sans groupement acétal. Le glucane est un résidu de glucoses formant des polyoses liés en β1–>3 et ramifié en β1–>6.F Croissance chez les champignons.
Il n’y a jamais de croissance intercalaire ==> Il y a toujours croissance apicale.L’apex contient l’activité mitotique du filament et possède des propriétés biologiques particulières. Le diamètre d’un filament est constant.
La paroi est rigide (indéformable) et il existe une polarité en direction de l’apex (mouvements de cytoplasme avec accroissement de la pression de turgescence).
G Organisation physiologique de l’apex.
1 : zone riche en vésicules ; 2 : une zone riche en mitochondries et REG et un certain nombre de filaments d’actine ; 3 : noyaux et vacuoles qui permettent d’apporter la pression de turgescence nécessaire à l’élongation.Suivant les types de champignons, l’agglomération des vacuoles sera différente : diffuse, à l’extrémité. Les ascomycètes présentent une densité aux extrémités (= spitzenkörper). Chez les basidiomycètes, les spitzenkörper ne sont que des vésicules en anneau autour. On a une accumulation de vésicules impliquées dans la synthèse de la nouvelle paroi au niveau de l’apex.
Les vésicules migrent vers l’apex par des microfilaments d’actine. Ces vésicules fusionnent ensuite avec la membrane plasmique et il y a extrusion de leur contenu (polymères de la paroi).
Au niveau du dôme, il y a expulsion de monomères de chitine, ce qui crée une zone élastique. Sur le côté, il y a libération de la chitine-synthase qui va polymériser la chitine. Cette enzyme est expulsée au niveau du dôme.
H Formation des ramifications.
Si le diamètre est constant, le thalle ne peut pas grossir, il va donc se ramifier pour « évacuer » l’excédent de nutriments. Après les premières cloisons, il y a formation des premières ramifications (indifféremment dans l’espace). Il faut qu’il y ait « dissolution » de la paroi pour pouvoir former un nouvel apex : nécessité d’enzymes lytiques. La ramification est aussi une alternative à la mobilité et à la dissémination. Ainsi, les colonies de champignons sont radiaires. Les parties vieilles sont au centre alors que les parties jeunes sont aux extrémités.I Formation de septums.
La formation des septums est centripète : elle commence au centre et s’étend vers les bords jusqu’à obturer les filaments.Les cloisons ont au milieu un pore permettant le passage du cytoplasme. Il sert à la communication des cellules et au passage des nutriments.
Quand il y a mort d’une cellule, il y a fermeture de ce pore afin d’isoler cette cellule.
II La multiplication végétative.
Un individu se développe identique à lui-même. On trouve :- Les bourgeonnements : levures.
- Fragmentation du thalle
- Formation de cellules spécialisées.
A Bourgeonnement.
Il y a formation d’un bourgeon. Quand la cellule est aussi grosse que la cellule mère, elle peut se séparer ou bien rester accolée. Si elle ne se sépare pas, il y aura formation d’une file de cellules appelées pseudomycélium.B Fragmentation du thalle.
Dans ce cas, les cellules formées sont des orthospores. Les cellules commencent par accumuler des réserves. Leur paroi se gélifie et les cellules se séparent du filament. Ces cellules deviennent des spores capables de générer un nouveau filament (géotricum). Il y aura formation de clamidospores qui ont un rôle de conservation. Leur paroi est épaisse et apparaît double ; leur cytoplasme est riche en réserves et parait coloré.C La formation de cellules spécialisées : sporulation vraie.
1 Les spores endogènes.
Ces spores apparaissent chez les oomycètes par un phénomène de polyplanétisme. La spore ne peut former directement un filament. La spore doit s’enkyster et donner une nouvelle spore flagellée (spore de second ordre). C’est le diplanétisme, avec deux spores nageuses. Ces spores peuvent germer un nouveau filament ou donner, après un nouvel enkystement, d’autres spores (de troisième ordre). Ces ré enkystements successifs procurent la capacité de germer. Les saprolégniales (inféodés à l’eau) voit leur phénomène de polyplanétisme s réduire au cours de l’évolution et de la diminution de leur dépendance à l’eau (aplanes).2 Spores endogènes non flagellées.
Les spores non flagellées sont produites par les sporocystes, eux-mêmes, portés par des sporocystophores. On a trois types de sporocystophores :- Type 1 : à columelle, contient de nombreuses spores.
- Type 2 : sans columelle, à nombre limité de spores.
- Type 3 : le sporocyste ne contient qu’une spore. La paroi du sporocyste se confond avec la paroi de la spore : sporocyste conidioïde.
3 Spores exogènes ou conidies.
Ces spores se forment directement sur les thalles (septés) : les spores sont inféodées au vent ou aux insectes. Elles peuvent se former en chaîne et donner un filament court qui émet des conidies les unes à la suite des autres. Ces spores peuvent aussi provenir de phialide. La phialide est une partie qui bourgeonne itérativement des conidies. La formation de conidies dans des phialides en goulot donne des endoconidies. Les conidiophores enfermés dans un stroma sont des pycnides. Au fur et à mesure de leur formation, ils seront libérés. Ce fonctionnement ne concerne que les ascomycètes.III La reproduction sexuée.
A Les générations morphologiques.
1 La multiplication végétative.
Le gamétophyte peut donner des gamètes haploïdes ou bien des spores équationnelles. Le méiosporophyte donne des méiospores ou des spores équationnelles.2 La reproduction sexuée.
Un cycle digénétique isomorphe présente deux générations identiques. Un cycle digénétique hétéromorphe présentera deux générations distinctes morphologiquement.a Cycle monogénétique haplophasique.
On observe un gamétophyte haploïde. Les gamètes vont fusionner pour donner un embryon qui subit immédiatement la méiose et donne quatre cellules (spores) qui pourront subir des phases de multiplication végétative.b Cycle monogénétique diplophasique.
Il y a formation d’un embryon par une fécondation. Celui-ci pourra subir deux phases de reproduction sexuée. Des cellules spécialisées vont subir la réduction chromatique et immédiatement fusionner (fécondation). On observe un méiosporophyte diploïde.c Cycle trigénétique haplodiplophasique.
La réduction chromatique donne des spores qui restent à l’état haploïde sans fusionner. Elles peuvent rester en l’état durant un long moment. Quand elles fusionnent, elles donnent une espèce diploïde. Dans la seconde partie du cycle, on peut observer des étapes de multiplication végétative. La troisième phase comprend la fin de l’individu diploïde qui subit la réduction chromatique. Suivant le moment de la méiose et de la fécondation, on distinguera des phases nucléaires.3 Exemples de cycles.
a Allomyces arbusculus.
La fécondation, une planogamie, donne un planozygote qui va se poser, puis germe. En germant, il y a formation d’un nématothalle qui libère directement des zoospores diploïdes. Indirectement, il peut y avoir libération de zoospores haploïdes par l’intermédiaire d’un sporokyste (avec réduction chromatique). Le cycle est isomorphe.b Saprolegnia monica.
La reproduction sexuée se fait par des sporocystes qui libèrent des zoospores flagellés (polyplanétisme). La fécondation est une oogamie siphonogame. Le thalle, diploïde, va former des organes de reproduction : les oocystes chez les femelles (contiennent des oosphères) ou des spermatocystes chez les mâles (fécondation par les noyaux). La fécondation donne des zoospores à 2N qui vont germer et donner un thalle diploïde.c Plasmopara viticola.
Dans le gamétophyte femelle, on ne trouve que des noyaux. Un seul oeuf est formé.d Mucor mucedo.
Les spores haploïdes, servant à la multiplication asexuée, sont disséminées par le vent.Quand il y a rencontre de deux gamétophytes compatibles, deux gamétocystes vont fusionner (caryogamie). On obtiendra alors, dans la zygospore, un seul noyau, diploïde, suspendu entre un suspenseur et le gamétocyste. La zygospore, par réduction chromatique, donne deux spores + et deux (haploïdes).
e Les Ascomycètes, Peziza.
Le thalle est haploïde, septé : c’est un mycélium primaire.L’organe femelle est un ascogone (une cellule renflée, terminée par un poil, le trichogyne). Les organes mâles sont les anthéridies (réalisent la fécondation par les noyaux). Une fois l’ascogone atteint, on ne voit pas de fusion des noyaux : il y a mise en place du syncarion. Il y a formation de dangeardies par le phénomène du crochet. L’asque est le méiosporocyste. Le cycle est digénétique haplodiplophasique. Le gamétophyte débute le cycle.
Pendant la formation des asques, il y a migration des noyaux vers le haut. A la base, il y a mise en place de vacuoles qui poussent les noyaux : il y a formation d’une vésicule ascale à la périphérie du plasmalemme. Après la mitose, cette vésicule (ascale), se fractionne pour donner la membrane plasmique de chaque future spore. Les ascospores sont endogènes. La libération de ces spores va se faire par éclatement de la structure de l’asque. La paroi est formée de deux tuniques : la tunique externe est l’exoascus (tunique hydrophobe, rigide), la tunique interne est l’endoascus (tunique mince, lipophile, souple). On trouvera des asques bituniqués (ils ont deux tuniques visibles après coloration) ou des asques unituniqués (les tuniques sont collées et l’on n’en voit qu’une).
Les différents mécanismes d’ouverture des asques sont :
- Jack in the box : Les vacuoles poussent et font craquer la tunique externe. La tunique interne va sortir. Ce moyen d’ouverture est typique des asques bituniqués.
- Eclatement.
- Ouverture par anneau.
- Ouverture par clapet.
f Les hémibasidiomycètes : Puccinia graminis.
Puccinia donne la rouille du blé.Le cycle est trigénétique. Le cycle commence avec un gamétophyte qui va infester l’épine vinette. On trouve un épithélium haploïde sur la face supérieure (gamétophytes mâles) ainsi que sur la face inférieure (gamétophytes femelles). Les spermaties (non flagellées, immobiles) sont libérées avec une goutte de miellat pour attirer les insectes. Il y a mise en place d’hyphes flexueux pour récupérer les spermaties (d’origine différente). Ces hyphes flexueux sont en contact avec des écidies (appareil femelle). Les spermaties se collent à l’hyphe. Le noyau mâle entre et descend vers le noyau femelle. Il va y avoir formation d’un dicaryon avec mitoses successives et bourgeonnement (les écidiospores). Ces écidiospores vont infecter un deuxième hôte (le blé) et former un mycélium dicaryontique. Au bout d’un moment, il va y avoir différenciation des urédosores qui vont émettre des urédospores. Ces urédospores sont des cellules rondes à paroi épaisse, avec deux noyaux, et reliées à la feuille par un pédicelle. Les urédosores sont des zones de regroupement des urédospores.
A la fin de l’été, le mycélium forme un nouveau type de spores : les téleutosores (taches noires) où l’on trouve des téleutospores. Ces spores sont pédicellées avec deux noyaux qui subissent la caryogamie. Au printemps, il y a émission d’un filament (boyau) où un noyau diploïde passe à l’intérieur. Il y a dans le boyau, formation de quatre gamètes haploïdes (méiosporocystes = basides). Chaque noyau est isolé par une cloison transversale. Chaque cellule à noyau haploïde bourgeonne une petite cellule où vient le noyau : les basidiospores. Ces dernières formes sont légères et vont réinfecter l’épine vinette.
La première génération est un mycélium haploïde. La seconde génération est composée des écidiospores et des urédospores. La troisième génération comprend les téleutospores et les basides (ou méiosporocystes).
g Les basidiomycètes, Coprinus comatus.
La fécondation est une somatogamie.Le mycélium primaire (haploïde) va faire une fécondation sans différenciation d’organes sexuels : il y a fusion de thalles complémentaires (plasmogamie). Il y a formation d’une cellule à dicaryon (pas de fusion). Il vient ensuite une anse d’anastomose (dangeardie). Il va y avoir une succession de cellules à dicaryon : le mycélium reste marqué par la bouche mycélienne, c’est le filament secondaire. C’est ce filament secondaire qui va former le carpophore. Au pied, il reste une membrane venant de la collerette qui se déchire : la volve. Au-dessus de cette volve, on trouve une cicatrice (un anneau).
Les lamelles sont des filaments dicaryontiques. Ce sont aussi des zones fertiles. Elles constituent l’hyménium :
- Cystites (réserves)
- Cellules sécrétrices (donnent du latex)
- Basides (cellules où a lieu la caryogamie).
C’est une basidiospore qui va germer un mycélium haploïde. Sur les quatre basidiospores on trouve deux cellules + et deux cellules -.
Chez les Auriculariales, le thalle est cloisonné. On a les basides en position latérale puis émission d’un long boyau d’où sortent les basidiospores. Les Urédinales et les Auriculariales ont des archéobasides. Les Tremellales ont des hétérobasides. Les Agaricales ont des basides typiques.
Les champignons montrent des formes très originales :
- On trouve des caryogamies différées.
- Il y a absence de gamètes mâles. Ils sont remplacés par des conidies (spores de multiplication végétative).
- Il peut y avoir autogamie, avec deux gamètes femelles.
- La somatogamie est une fusion de deux thalles qui donne un mycélium à dicaryon.
IV L’hétérotrophie.
A Hétérotrophie vis à vis de l’azote.
Parmi les autotrophes, on peut trouver des levures, qui, dans certaines conditions, vont pouvoir utiliser l’azote atmosphérique. Des mycètes comme Aspergillus niger et Mucor mucedo vont absorber de l’azote nitrique. D’autres individus sont appelés semi-hétérotrophes (Mucor sp., des levures) et sont caractérisés par une consommation d’azote ammoniacal.Les hétérotrophes peuvent être des saprophytes comme Saprolegnia, des symbiotiques comme les champignons lichénisants, des parasites comme Phytophtora, Plasmopara ou bien des prédateurs comme Dactylella acrochaete. Ces individus consomment de l’azote organique.
B L’hétérotrophie vis à vis du carbone.
1 Le saprophytisme.
On peut trouver trois types de pourritures :- Pourriture cubique.
- Pourriture fibreuse.
- Pourriture alvéolaire.
2 Le parasitisme.
Comme type de parasitisme, on peut citer la graphiose de l’orme (maladie de l’orme champêtre), la maladie de la pomme de terre irlandaise.Ce parasitisme peut s’effectuer par une effraction tissulaire et même parfois, cellulaire (avec la formation de suçoirs). Elle peut aussi se présenter par une action biochimique avec la sécrétion d’enzymes, de toxines, de substance de croissance. Ces actions provoquent un flétrissement (les toxines) ou un gigantisme (substances de croissance).
On trouve des champignons parasites de faiblesse.
Ces champignons se développent sur une plante même quand elle est morte : ce sont des nécrotrophes ou bien saprophytes. Il existe différents types de biotrophie avec aussi des développements sur des plantes vivantes. Ce sont des parasites forts ou obligatoires.
Les différentes étapes nécessaires à une infection :
- Reconnaissance de la plante par échange de signaux biochimiques. On trouve des spectres étroits ou bien plus larges. (spectre étroit pour les parasites forts et large pour les parasites faibles).
- Attachement à la surface de la plante.
- Effraction tissulaire.
- Développement interne, avec effraction cellulaire possible.
- Troubles chez la plante.
- Reproduction du champignon.
a L’adhésion.
Cette adhésion se fait par sécrétion d’un mucilage de polysaccharides et de glycoprotéines sous le tube germinatif. Il va y avoir synthèsed’enzymes pour dégrader l’épiderme de la plante.
b La pénétration.
La pénétration peut se faire par voies naturelles (comme les stomates) et dans ce cas, il n’y a pas de différenciation d’un tube germinatif, au niveau de l’épiderme ou des racines, ce qui est une pénétration directe. Dans ce dernier cas, il va y avoir mise en place d’organes particuliers (actions mécaniques) et libération d’enzymes pour dégrader la paroi (actions chimiques).Pour l’action mécanique, il va y avoir renflement du tube germinatif (ou appressorium) : la pression osmotique du champignon est plus forte que celle de la plante afin d’y résister.
Il va ensuite y avoir formation d’un filament fin (point d’infection) duquel partent des filaments infectieux qui portent les suçoirs (Haustorium).
Ce sont les structures infectieuses.
Les champignons à appressorium mélanisés vont renter sans problème. Les formes à appressorium muté ne rentrent pas. La mélanine permet l’accroissement de la pression pour la pénétration.
Lors de l’action chimique, la formation du tube germinatif entraîne la formation d’un mucilage, d’enzymes hydrolithiques pour la dégradation de l’épiderme de la plante (cutinases, lipases, cellulases&) Il va y avoir une diminution de la paroi du végétal.
La pénétration mécanique par les racines. Deux cas existent pour cette pénétration : coussins d’infection et rhizomorphes. Les coussins d’infections sont des agglomérations d’hyphes d’où partent plusieurs points d’infection (les coins d’infections). Ils partent en parallèle et attaquent la plante. Les rhizomorphes sont des extensions gélatineuses qui permettent l’adhérence. Ce sont des émissions latérales qui pénètrent car ce sont des filaments agglomérés.
Une fois à l’intérieur, deux cas sont possibles :
- Nécrotrophe : il y a dégradation des parois du végétal puis des parois cellulaires. Les filaments s’insinuent dans toutes les fentes mais provoquent aussi la mort de la plante.
- Biotrophe : Les filaments migrent dans la plante, au niveau de la lamelle moyenne. Seuls les suçoirs entrent dans les cellules. Ces derniers permettent aussi la fixation.
Les toxines vont provoquer un flétrissement de la plante. Les substances de croissance, provoquent elles, un gigantisme mais aussi un nanisme et une castration sont possibles.
3 L’endophytisme.
L’endophytisme est entre le parasitisme et la symbiose. C’est un cas qui provoque peu de dégâts.a Chez Claviceps.
L’infestation ne provoque aucun symptôme, sauf à la floraison où dans l’ovaire, le stroma donne des conidies. Le stroma va se différencier en sclérote qui tombe au sol. Là, il se développe des structures de reproduction sexuée.b Chez Epichlöe triphina (la quenouille de graminées).
Il va se former un stroma blanc autour de la tige. C’est là où sont les structures de reproduction.c Chez Sphacelia tryphina.
Ce champignon est responsable de l’ivraie enivrante. Il se développe des graines qui donneront une plantule avec le champignon. Les nouvelles graines porteront toujours le champignon. Les animaux qui consomment ces graminées auront des malaises et autres symptômes similaires car ces champignons synthétisent des alcaloïdes.4 Les symbioses.
On distinguera deux cas de symbioses : la symbiose par les mycorhizes ou par les lichens. Les mycorhizes se trouvent chez les ascomycètes, les basidiomycètes ou chez les zygomycètes (Glomales, la reproduction sexuée est diminuée).Dans le cas de l’ectomycorhize, le champignon se développe à l’extérieur ; il ne se trouve jamais dans les cellules ; il provoque des modifications morphologiques des racines ; il se développe autour des racines.
Dans le cas de l’endomycorhize, le champignon ne montre pas de développement important à l’extérieur ; il entre à l’intérieur des cellules ; il n’entraîne pas de modifications importantes des racines.
a L’endomycorhize (Les Glomales).
Les glomales sont uniquement symbiotiques. On trouve quatre familles : Glomus, Gigaspora, Sclérocystis, Acannospora.Une spore va germer et donner, à l’extérieur, un filament et un appressorium (comme chez les parasites). Il se forme des arbuscules dans les cellules végétales (suçoirs). Dans certaines familles comme Glomus, il y a formation de grosses vésicules contenant des réserves pour le champignon : les méats. Les arbuscules sot des surfaces d’échange entre le champignon et la plante (Gigaspora). La plante va former une matrice extra-fibrillaire pour empêcher la prolifération des arbuscules (durée de vie : 3-4 jours).
Les mycorhizes d’Ericaceae. La racine est fine à petit diamètre et simple (cortex, endoderme, pôle ligneux). La colonisation du champignon ne se fait que dans les cellules corticales. Les racines mycorhizées vont dégénérer au fur et à mesure.
b L’ectomycorhize.
Cette forme de symbiose fait intervenir les basidiomycètes et les ascomycètes (Tuber melanosporum). Il va y avoir formation d’un réseau extra-matriciel (mycélium dans le sol) et d’un réseau interne (réseau de Hartig). Il n’y a pas de pénétration intracellulaire (reste dans les méats). La racine va se transformer. Le réseau de Hartig est une surface d’échange entre le champignon et la plante.c L’ectendomycorhize.
Les mycorhizes à Orchidée.Un basidiomycète dont la forme végétative est Rhizoctonia va coloniser les racines adultes et l’embryon (dépourvu de réserves). La colonisation de l’embryon permet sa germination. Il faut un équilibre entre la phagocytose et le parasitisme pour que l’embryon puisse atteindre la germination.
C’est un état de parasitisme contrôlé pour une vie commune. Cette ectendomycorhize est un type intermédiaire. Il y a formation d’un manchon et une pénétration intracellulaire.
d Conclusion sur les symbioses.
La symbiose permet aux champignons d’obtenir des synthétats, source de carbone. La plante, grâce au champignon, peut aller chercher l’eau et les sels minéraux (P, Cu, Z, S) beaucoup plus loin. Certains champignons vont même jusqu’à récupérer des ions comme NO3- et bien d’autres pour que la plante puisse se développer sur de mauvais milieux. Il y a aussi l’action des hormones de croissance. Les Herbacées sont endomycorhizables alors que les Arbres sont ectomycorhizables.5 La place des champignons dans la biosphère.
Les saprophytes permettent la dégradation des plantes mortes et vont former l’humus et donc, améliorer les sols.L’endomycorhize permet l’accroissement de l’absorption de l’eau et des sels minéraux. Les cellules lignivores participent à la nutrition des herbivores. Les parasites participent à l’équilibre des populations végétales.V La symbiose lichénique.
Un lichen est une association stable entre une algue et un champignon qui donne au final, un nouvel individu. Les lichens sont des organismes polyphylétiques. Ils sont apparus à 5 reprises dans l’évolution. Les plus vieilles formes ont environ 400 millions d’années (au Dévonien). On en connaît 25 000 espèces.Le partenaire fungique est appelé « mycobionte ». Ce sont essentiellement des ascomycètes (et souvent des Pyrénomycètes et des Discomycètes). On trouve tout de même 1% de basidiomycètes. 13 250 espèces de champignons sont lichénisantes.
Chez les algues, photobiontes, on trouve 40 espèces lichénisantes. Exemple : Trebouxia, Pseudotrebouxia, Trentepholia, Nostoc.
A Les diverses formes de lichens.
1 Les lichens gélatineux.
Ces lichens sont homomères. Les hyphes sont mélangés aux cellules des algues. Ils sont homogènes dans le désordre (en général, Ascomycètes + Nostoc).2 Les lichens secs.
Ces lichens sont toujours organisés en strates. Ils sont appelés hétéromères (structure hétéromère stratifiée sauf chez les lichens fructiculeux). Parmi eux, on trouve :- Les lichens secs crustacés : Ces lichens sont sur les rochers (rupicoles), les trottoirs. Ils sont complètement dans leur substrat. De l’extérieur (air) vers l’intérieur (contre le substrat), on trouve : un mycélium (le cortex), une couche gonidiale et une médulla arachnoïde.
- Les lichens secs foliacés : Ces lichens sont sur toute la longueur du substrat. De l’extérieur vers l’intérieur, on trouve : un cortex supérieur, une couche gonidiale (cellules algales), une médulla arachnoïde et un cortex inférieur avec des rhizines.
- Les lichens secs fructiculeux : Ces lichen fructiculeux pendent des branches d’arbres. Ils ne sont appliqués au substrat que par un point. Ils ont, en coupe, une forme relativement cylindrique. L’extérieur vers le centre, on trouve : un cortex supérieur, une couche gonidiale (avec cellules algales), une médulla arachnoïde puis une chondroïde. Ces lichens ont une structure radiée.
3 Le thalle secondaire.
Ce thalle secondaire est appelé podétion. En coupe, un podétion présente : un cortex supérieur, une couche gonidiale, une médulla arachnoïde creuse, une médulla chondroïde et une lacune centrale.Les cyphelles sont des trous pour l’aération de la couche gonidiale.
Les céphalodies apparaissent quand deux lichens sont superposés. Un champignon, une chlorophycée et une cyanobactérie. On a donc un thalle à cyanobactérie et un thalle à chlorophycée ainsi qu’un double champignon. Il peut y avoir fixation d’azote atmosphérique grâce aux cyanobactéries.
B Les structures de multiplication et reproduction.
Les pycnides. Ce sont des structures de multiplication du champignon qui forment et libèrent des conidies.Les isidies. Ce sont des structures dressées, ramifiées à la surface du thalle. Elles forment de petits cylindres constitués d’algues, entourés de filaments mycéliens. Elles sont détachables au vent.
Les sorédies. Les sorédies sont des cellules d’algues individuelles, entourée d’hyphes regroupés dans une soratie.
Le champignon, seul, possède des structures d reproduction :
- Des apothécies si ce champignon est un discomycète. Elles sont sur les bords du thalle et forment un petit sillon convexe ou concave.
- Des périthèces si le champignon est un pyrénomycète. Elles prennent la forme d’une urne, seule, ou en groupe (se trouvent surtout chez les
lichens crustacés).
Croissance et longévité.
Les crustacés ont une croissance de 2 à 3 millimètres par an. Les autres lichens ont une croissance comprise entre 3 et 4 centimètres par an.La croissance est composée de plusieurs phases :
- Phase I : croissance optimale.
- Phase II : phase de maturation.
- Phase III : phase de dégénérescence (au centre) alors que l’on observe une croissance de l’extérieur.
C La symbiose lichénique.
1 Symbiose contre parasitisme.
Le champignon absorbe les photosynthétats de l’algue. Celui-ci amène l’eau, protège de la dessiccation, des UV et du broutage par les herbivores grâce aux acides lichéniques.2 Le parasitisme.
Dans ce cas, c’est le champignon qui va parasiter l’algue : on parle d’hélotisme. L’algue est prisonnière dans le champignon. Il y a inhibition de sa reproduction sexuée et sa photosynthèse est modifiée en quantité et en qualité.3 L’algue parasite le champignon.
Les céphalodies sont considérées, par certains, comme des galles. On observe des réactions de défense du champignon contre l’algue.- En fait, on observe un état d’équilibre et des relations mutualistes.
- L’antagonisme du champignon est compensé.
- Il y a synthèse d’un nouvel individu : le LICHEN.
- La somme du champignon et de l’algue fait apparaître des propriétés propres aux lichens.
4 Relation morphologique entre algue et champignon.
On distingue trois types :- Cohabitation sans contact entre les cellules de l’algue et celles du champignon.
- Le champignon enserre l’algue. Pour cela, il va former des crampons.
- Haustorium. Si l’algue est une cyanobactérie, le champignon peut entrer dans le mucilage, mais pas dans les cellules. Si l’algue est une chlorophycée, le champignon peut faire un Haustorium qui entrera dans la chlorophycée.
5 Relations biochimiques.
Les algues fournissent les photosynthétats (1/3 dans le cortex et 2/3 dans la médulla).Quand le champignon récupère les sucres, il va les transformer en sucre-alcool (ribitol, mannitol…). Ces sucres modifiés permettent le maintien de la turgescence du lichen et le protège d’une dessiccation trop élevée.
Les champignons vont synthétiser des métabolites secondaires (antibiotiques et acides lichéniques). Ces acides vont intervenir dans la régulation du nombre de cellules d’algue (en le faisant diminuer). Toutefois, ils protègent aussi les algues contre les UV et contre la dessiccation. Les champignons interviennent aussi dans la protection contre les métaux lourds.
La classification est basée sur la classification des champignons, puis sur les modes de reproduction.
D Les lichens dans leur milieu.
Les lichens ont une teneur en eau très faible (2 à 10% du poids sec du lichen). S’ils se dessèchent, il y a diminution de la photosynthèse. Le cortex va alors s’épaissir, devenir opaque et l’algue ne reçoit plus d’UV. Ils peuvent toutefois se réhydrater très vite : poïkilohydre. Il y aura alors reprises des fonctions métaboliques. Le cytoplasme doit être résistant.Il n’y a photosynthèse que durant les premières heures d’ensoleillement. Dès que le climat devient trop sec, il y a arrêt de la photosynthèse. Ce qui explique la lente croissance des lichens.
Les lichens sont quand même capables de coloniser des milieux extrêmes : on les trouve dans la toundra, dans l’antarctique (350 espèces de lichens contre deux espèces deux plantes vasculaires): ce sont des pionniers.
Les facteurs écologiques pour l’installation des lichens :
- Types de substrats (milieu, composition&)
- Climats (humidité, ensoleillement)
- Facteurs biologiques : animaux intervenant sur leur installation (déjection) et pollution humaine.
1 Les lichens terricoles.
Ces lichens sont les plus soumis à des modifications de substrat. Ils sont sur l’humus, le calcaire, avec des végétaux, avec des débris organiques.Si l’on se trouve dans des prairies semi-désertiques, on a une espèce de Pormélia (foliacée) qui va donner une espèce de Cladonia puis disparaître. Cladonia (Podétiée) donne à son tour 4 à 5 espèces nouvelles de Cladonia mais l’espèce d’origine disparaît. Sur ces 4 ou 5 espèces, 3 ou 4 vont disparaître et une survivra.
2 Les lichens épiphytes.
On trouve des successions lichéniques différentes si le végétal est isolé ou en groupe (forêt). L’espèce du végétal colonisé intervient aussi. Les écorces lisses à pH alcalin hébergeront plutôt des lichens foliacés alors que les écorces rugueuses, à pH acide, hébergeront des lichens fructiculeux.3 Les lichens lithicoles.
On trouve différents lichens selon la nature chimique. A pH basique, on aura un lichen comme Collema alors qu’à pH acide on trouvera des crustacés. On distinguera aussi des lichens calcicoles et des lichens calcifuges. La nature chimique pourra être modifiée par l’humus, les déjections d’oiseaux (chargées en azote). La nature physique pourra être modifiée par les caractères des failles.4 Les lichens aquatiques.
α Les lichens marins.
Ils sont sur les roches acides et en milieu marin. Ils sont différents en fonction des zones de balancement des marées. Au niveau de la zone intertidale, on trouve Verrucaria qui forme une ligne noire. Lichina forme une ligne brune avec les embruns, les marées, les équinoxes. Calauplaca et Xanthoria forment une ligne orange.β Les lichens d’eau douce.
Il existe des lichens émergés 6 mois par an, d’autres émergés seulement 3 mois par an et d’autres qui ne sont jamais émergés.Les lichens n’ont pas le même système de filtration : pour cela, il y a utilisation des acides lichéniques.
Les pollutions par H2SO4 (ou d’autres polluants) peuvent tuer les lichens.