
I Protothériens ou Monotrèmes (mammifères inférieurs).
Exemple : l’ornithorynque.Il y a des ovocytes riches en réserves, s’entourant d’albumen au niveau de l’oviducte puis d’une coque cornée contenant des nodules calcaires. Seule l’ovaire gauche est fonctionnel. L’œuf est pondu par la femelle dans un terrier.
Chez les échinides (ils ressemblent à des petits hérissons et sont proches des marsupiaux), il y a incubation des œufs dans une poche ventrale abdominale dans laquelle ils sont reçus au sortir du cloaque. Les deux ovaires sont fonctionnels et l’œuf pondu correspond à un embryon de poulet à 40 heures d’incubation.
On distingue trois périodes chez ces espèces :
- Une période intra-utérine (brève).
- Une période d’incubation.
- Une période qui suit l’éclosion et où commence la lactation.
II Les métathériens (les marsupiaux).
La gestation est de courte durée, l’embryon se développe dans l’utérus où il effectue son organogenèse. Il y a formation d’un placenta très rudimentaire. Quand le jeune est mis au monde, il gagne la poche marsupiale où il achèvera sa croissance.III Les euthériens.
La gestation va atteindre son maximum de compétence. L’œuf est dépourvu de réserve (alécithes). Le développement va dépendre de substances nutritives apportées par la mère grâce à un organe d’origine fœto-maternelle qu’est le placenta.IV Physiologie de la gestation.
L’œuf ne peut s’implanter dans la muqueuse utérine que si celle-ci est préalablement préparée par l’action synergique des œstrogènes puis de la progestérone.L’ovariectomie précoce empêche la nidation. Or, quand la gestation est installée, on distingue deux types d’espèces :
- Espèce où l’ovaire est indispensable jusqu’au terme (exemple : le lapin, le hamster).
- Espèce où l’ovaire n’est plus nécessaire dans la deuxième partie de la grossesse (femme, cobaye, chien, chat) : le placenta prend le relais hormonal et sécrète une gonadotrophine agissant sur le corps jaune cyclique qui devient ainsi gestatif. La grossesse se résume par un équilibre entre œstrogènes et progestérone, sans oublier les interactions hypophyse, hypothalamus.
V L’ovoimplantation.
Les œufs alécithes (de 100 à 150 µm de diamètre).Dans les douze heures qui suivent la ponte, l’ovocyte II doit être fécondé, sinon il dégénère. La première mitose a lieu vers la trentième heure. La segmentation est asynchrone. Au bout de quatre jours, on obtient un stade morula, formé de douze à seize cellules. Le transit tubaire est favorisé par un flux péritonéo-utérin en péristaltisme de l’oviducte et de l’action des cils.
La morula arrive dans l’utérus où elle atteint un stade blastocyste (vers le 6ème jour). Ce blastocyste est formé de deux parties :
- des cellules aplaties à la périphérie
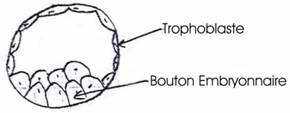 un amas cellulaire à l’un des pôles constituant le bouton embryonnaire.
un amas cellulaire à l’un des pôles constituant le bouton embryonnaire.
VI Les différents types de nidation.
A L’implantation centrale.
Dans ce cas, la muqueuse utérine est peu transformée. Le blastocyste reste dans la lumière de l’utérus et s’y développe. Ses parois viennent alors s’accoler à celles de l’utérus. L’endomètre est peu transformé. C’est un phénomène que l’on observe chez les carnivores, les ruminants, les suidés et les chéiroptères.B L’implantation excentrée.
Le blastocyste s’insinue entre les plis de la muqueuse utérine qui est fortement transformée et qui ressemble à une véritable dentelle. La transformation de l’endomètre se fera grâce à des hormones. Le blastocyste détruit partiellement la muqueuse utérine, se refermant sur lui-même pour former une chambre close, sans communication avec la lumière utérine. Cette implantation a lieu chez le lapin, le rat et la souris.C L’implantation interstitielle.
Le blastocyste pénètre dans l’épaisseur de l’endomètre et s’y développe. C’est le cas des primates, de l’homme. La grossesse ectopique (extra-utérine) est fréquente chez la femelle, dans la trompe, l’abdomen ou sur un ovaire.VII La placentation.
Le placenta s’édifie à partir du trophoblaste, du blastocyste et de la muqueuse utérine.A Formation du côté fœtal.
La prolifération cellulaire se poursuit activement au niveau du trophoblaste. Au 7ème jour, chez la femme, le trophoblaste se différencie en un cytotrophoblaste cellulaire et à sa périphérie, on a un syncytiotrophoblaste qui, par son activité protéolytique, attaque l’épithélium, le tissu conjonctif et la paroi endothéliale des vaisseaux maternels. Cette nidation est complète au bout de douze à quatorze jours.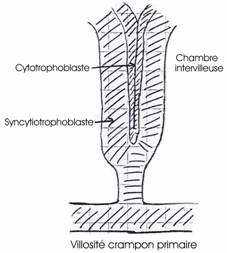
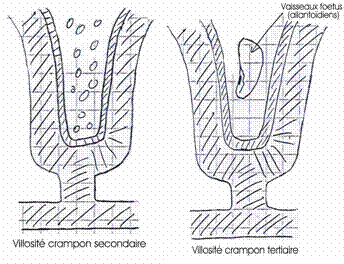
Le syncytiotrophoblaste constitue des chambres intervitellines pleines de sang maternel séparées par des évaginations du cytotrophoblaste (ou villosité crampon primaire) avec un axe mésodermique (villosité crampon secondaire), puis, par la présence de vaisseaux fœtaux (villosité crampon tertiaire).
B Formation du côté maternel.
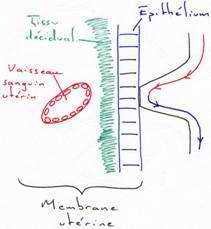
Sous l’effet des hormones, l’endomètre utérin se modifie : c’est la réaction déciduale qui est plus ou moins importante selon les espèces. L’endomètre prend le nom de décidu ou de caduque qui joue un rôle protecteur contre les attaques immunologiques de la mère et contre l’activité érosive du syncytiotrophoblaste. La décidu peut se conserver intacte au moment de la mise bas ou parturition (accouchement chez la femme).L’expulsion du placenta se fera sans hémorragie maternelle : on parle alors de placenta indécidué (sans caduque). La décidu est partiellement détruite par le syncytiotrophoblaste. Il y aura élimination de la décidu à la naissance : on parle de placenta décidué.
C Formation de l’allantoïde et évolution de la placentation.
Chez les oiseaux, l’allantoïde est une invagination de l’intestin postérieure doublée extérieurement de splanchnopleure très vascularisée. Elle se forme tardivement chez les oiseaux (vers 60 heures d’incubation), quand la moins grande partie des somites est en place. Chez les mammifères, sa formation est beaucoup plus précoce, avant même l’apparition des premiers somites, vers le 18ème jour chez la femme.Il y a deux annexes :
- La vésicule vitelline est à l’origine d’un amphaloplacenta.
- L’allantoïde donne un allantoplacenta.
L’évolution du placenta se fait par une réduction progressive de la vésicule vitelline et le développement de l’allantoïde, puis, chez les mammifères supérieurs (primates et hommes), elle se fait par une réduction d’allantoïde. A partir du troisième mois, chez la femme, la vésicule vitelline et l’allantoïde dégénère et à la naissance persistent deux artères et un ucine.
Chez les marsupiaux, le placenta est très rudimentaire et aussi bien constitué par la vésicule vitelline que par l’allantoïde. Chez les péramèles (marsupiaux), il y a une volumineuse vésicule vitelline et un allantoïde qui vient s’appliquer contre le chorion. Il se forme ainsi un alloplacenta sans villosité. Le fœtus dépend de la sécrétion vitelline (une sorte de lait utérin) pour son développement. Il y a une vascularisation de la vésicule vitelline (chez les marsupiaux). La circulation allantoïdienne apporte au fœtus des substances nutritives.
Chez les carnivores, la vésicule vitelline est nettement réduite mais persiste durant la gestation, avec sa vascularisation, mais elle n’atteint pas le chorion. L’allantoïde s’étale sur toute la face intérieure et entoure la cavité amniotique : on a alors un placenta dit allantoïdien. Les villosités se localisent dans une zone, d’où le nom de placenta zonaire.
Chez les ruminants, l’allantoïde est extrêmement développé ainsi que l’amnios. Chaque fois que l’allantoïde régresse, les vaisseaux allantoïdiens s’étendent dans la membrane chorio-amniotique, le placenta restant allantoïdien.
1 Les différents types d’alloplacenta.
Les alloplacentas sont classés en deux catégories :- selon la typographie des villosités choriales,
- selon leur relation avec l’endomètre utérin.
2 le placenta épithéliochorial :
L’épithélium de l’endomètre vient en contact avec les villosités choriales. On en distingue deux types :- le placenta diffus. Dans ce cas, les villosités choriales sont réparties de façon uniforme, elles s’enfoncent dans les glandes de l’endomètre remplies d’embryotrophe (produit de sécrétion absorbé par les villosités choriales). C’est le plus primitif. Il est observé chez les marsupiaux.
- Le placenta indécidué. On le trouve chez les cétacés, les chevaux, les porcs.
3 Le placenta cotylédonaire.
Dans ce cas, les villosités choriales s’enferment dans un épaississement de l’endomètre en caroncule. L’ensemble des villosités choriales et de la caroncule constitue le cotylédon.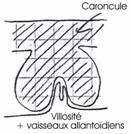
4 Le placenta endothéliochorial.
Le syncytiotrophoblaste vient en contact avec l’endothélium des vaisseaux de la mère (chez les carnivores). Le placenta est zonaire et décidué.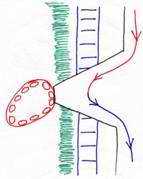
5 Le placenta hémochorial.
Les villosités choriales érodent l’épithélium des vaisseaux de la mère, provoquant une hémorragie. Il y a formation de lacunes de sang maternel (comme chez la femme). C’est une placentation discoïdale).D Le rôle physiologique des placentas.
La surface des villosités est de l’ordre de 14 m². Les échanges se font parDiffusion simple (pour l’O2, le CO2, l’eau, le Na+, le K+, les chlorures et les bicarbonates).
Diffusion facilitée : les molécules sont transportées d’un pôle de la cellule à l’autre (ex : les sucres, les sels, les acides aminés …).
Transport actif : il est nécessaire pour les molécules plus complexes.
La phagocytose et la pinocytose.
Les substances transportées par le sang maternel passent dans le sang fœtal (les gaz, l’eau, les électrolytes, les glucides, lipides, protides, vitamines, hormones et anticorps).
Chez l’homme, l’immunité passive est transmise in utero. Beaucoup de médicaments traversent, ainsi que pratiquement tous les antibiotiques. Ces substances peuvent être accompagnées, dans leur traversée, par des agents infectieux et par des virus.
E La fonction endocrine du placenta.
Le placenta sécrète une HCG : Hormone Gonadotrophine Chorionique. Elle est produite par le syncytiotrophoblaste, éliminé dans l’urine de la femme enceinte. L’injection à des lapins impubères provoque le maintien du corps jaune (de cyclique, il devient gestatif).L’HLP est l’Hormone Lactogène Placentaire. Elle provoque la sécrétion lactée chez les lapines,… Elle est analogue à la prolactine. Elle n’a pas un rôle lactogène mais participe avec les stéroïdes sexuels dans la préparation de la glande mammaire à la lactation. Cette hormone contrôle la mobilisation des lipides maternels.
L’œstrogène et la progestérone agissent sur l’utérus : il y a stimulation de la croissance et du métabolisme. Elles agissent sur le fonctionnement des systèmes enzymatiques placentaires. Dans les deux premiers mois, la progestérone vient du corps jaune, puis, du placenta. Son effet majeur est l’inhibition du muscle utérin.