I Les œufs.
Comme chez tous les vertébrés, l’organogenèse est un phénomène discontinue. Après une phase de multiplication, les ovogonies deviennent des ovocytes primaires restant bloqués en prophase méiotique. Viennent ensuite les ovocytes secondaires qui, une fois libérés de l’ovule, sont captés par l’oviducte. Il y a lors fécondation. La rupture de l’ovaire et la fécondation, régulent l’ovogenèse.En trois ans, l’ovocyte primaire augmente de taille : elle passe de cinquante micromètres à 2 millimètres. Ces ovocytes permettent le stockage de matériaux dans les ovaires. L
es matériaux stockés sont de 2 types :
- nutritifs (vitellus).
- endogènes (ARN messagers, transcrits), ils permettent une multiplication nucléaire. Ils permettent aussi la transition blastuléenne par une traduction en protéines.
Dans l’ovaire, l’ovocyte primaire augmente considérablement de taille durant trois ans chez Rana esculenta, avant la ponte ovulaire. Le noyau constitue la vésicule germinative : le noyau est bloqué en prophase 1 de méiose.
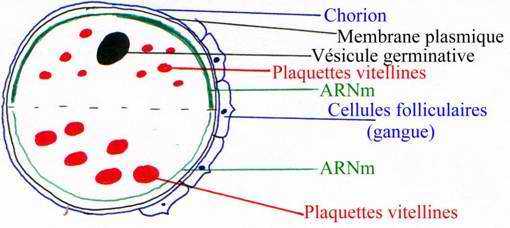
Dans l’ovocyte primaire, il y a stockage de matériaux nutritionnels d’origine exogène apportés par la circulation maternelle. Il y a des protéines, des vitellogénines qui sont synthétisées par le foie de la mère et captées par les cellules folliculaires entourant l’ovocyte puis, transformées par l’ovocyte après déshydratation en complexes phosphoglycolipoprotéiques. Les phosphoglycolipoprotéines sont appelées plaquettes vitellines.
L’ovocyte primaire stocke aussi du matériel d’origine endogène permettant l’expression de l’information génétique. Il s’agit de ribosomes : on parle d’amplification de cistrons ribosomiques.
A maturité, cet ovocyte primaire a un noyau volumineux excentré vers le côté. Il se différencie en 2 hémisphères : un pigmenté qui représente le pôle animal et un autre, dépigmenté qui représente le pôle végétatif.

L’ovocyte va stocker des matériaux exogènes. Le précurseur est la vitellogénine (470 kDa). Celle-ci est synthétisée par le foie de la femelle, elle circule dans le sang et va être internalisée dans des endosomes puis dans des lysosomes et, elle sera enfin dégradée en phosphovitine et lipovitelline qui vont être déshydratées et qui vont donner les plaquettes vitellines (structure cristalline). Elles sont stockées en réserve puis dégradées par des enzymes le moment venu. Au pôle végétatif : les grosses plaquettes vitellines vont rester sur place. Au pôle animal, les petites plaquettes vitellines vont être transportées par des filaments d’actine vers le centre de la cellule : on obtient un gradient vitellin. On a alors une distribution symétrique des molécules.
Les matériaux endogènes : ont trouve du glycogène stocké autour des noyaux, des ribosomes, du réticulum endoplasmique et des lipides (tous sont répartis autour du noyau). On trouve de l’ARN de tous types qui est réparti d’une certaine manière. L’ARN est plus riche vers le pôle animal que vers le pôle ventral.
L’ARN messager Vg va coder pour la synthèse de facteurs de développement du pôle ventral dans l’induction du mésoderme. D’autres ARNm vont coder pour la synthèse de molécules adhésives, de catéine qui entraîne une localisation préférentielle des ARN au pôle animal ou ventral.
Sous la membrane plasmique, il y a des granules pigmentaires qui confèrent une couleur à l’œuf : on parle de calotte pigmentaire dans le cytoplasme cortical qui contient beaucoup de vitellus, de mitochondries, de granules corticaux.
La taille de l’ovocyte I est comprise entre 5 µm et 2 mm.
Cet ovocyte se libère de l’ovaire et sera émis dans l’oviducte femelle. Il y a alors reprise de la première division méiotique, métaphase I, avec production d’un globule polaire qui donnera un ovocyte II qui restera en métaphase II dans l’oviducte. Il sera ensuite émis dans l’eau puis, fécondé par le mâle.
Au moment où l’ovocyte I est émis, il achève sa première (sinèse) division méiotique et rejette le globule polaire qui se manifeste au pôle animal par une région dépigmentée : la tache de maturation.
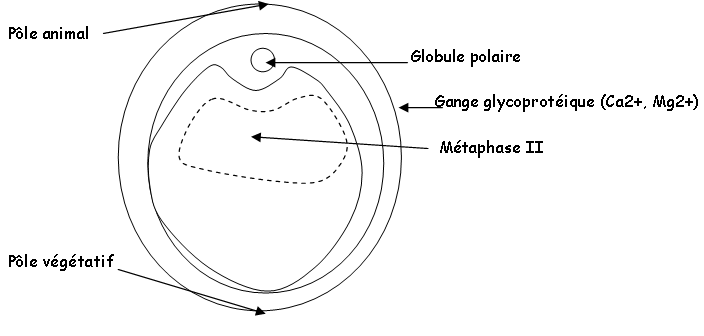
Cet ovocyte s’entoure d’une gangue glycoprotéique (muqueuse) qui est sécrétée durant le transport dans l’oviducte. Elle a pour fonction de créer des conditions favorables pour la fécondation. Elle apporte des ions comme Ca2+ et Mg2+. Elle a aussi un rôle protecteur en empêchant les œufs de se déshydrater quand ils sont émis hors du cloaque. Cette gangue fait adhérer les cellules entre elles : ensemble cohésif.
II La fécondation.
Celle-ci est variable chez les amphibiens. On distingue deux types distincts : la fécondation des anoures et des urodèles.A Les anoures. Exemple : le Xénope.
La fécondation est externe. Le mâle va chevaucher la femelle en répandant son sperme au moment où les œufs sont pondus.La fécondation doit avoir lieu dès l’émission des ovocytes par la femelle. La gangue, hydrophile, gonfle au contact de l’eau et devient rapidement un obstacle à la pénétration des spermatozoïdes (fécondation).
B Les urodèles. Exemple le Triton (pleurodèle).
La fécondation a lieu au niveau du cloaque de la femelle ; il n’y a pas d’organe copulateur. Le mâle émet un spermatophore (formation mucilagineuse) au niveau duquel sont agglutinés les spermatozoïdes. Ce spermatophore va adhérer au substrat (caillou).La fécondation est dite interne et se produit au niveau du cloaque (canal mixte urogénital et dont les lèvres viennent enserrer le spermatophore).
La pénétration des spermatozoïdes a lieu généralement au niveau du pôle animal car c’est là où il y a le moins d’encombrement.
Il y a monospermie chez les anoures (un seul spermatozoïde rentre) mais polyspermie chez les urodèles (5 ou 6 spermatozoïdes rentrent mais un seul va féconder le noyau). La fécondation va entraîner une activation dans l’ovocyte : deux types d’activation (interne et externe).
Dans le cas d’une activation interne, il y a reprise de la deuxième division méiotique avec émission d’un second globule polaire et la formation d’un pronucléus femelle.
Le spermatozoïde perd son flagelle. L’acrosome vient se fixer sur un récepteur, traverse la membrane vitelline et, au contact de la membrane plasmique, va produire une dépolarisation de la membrane en modifiant le potentiel de celle-ci. Cette dépolarisation de quelques minutes est suivie d’une repolarisation. Ce phénomène entraîne une libération de Ca2+ (Ca qui provient du réticulum endoplasmique lisse et des mitochondries) dans la région corticale de l’œuf. Ce Ca permet la polymérisation des filaments d’actine (allongement de la cellule), ce qui a pour effet d’amener à la surface des granules corticaux. Ces granules libèrent leur contenu en faisant fusionner leur membrane (exocytose). Ces granules corticaux contiennent trois composants :
- mucopolysaccharides qui vont être émis entre la membrane plasmique et la membrane de fécondation (vitelline), constituant ainsi un gel hydrophile amenant de l’eau entre ces membranes et les décollant.
- participant à la désolidarisation des 2 membranes.
- >Glycoprotéines épaississant la membrane vitelline (de fécondation) qui se décolle.
Remarque : les granules corticaux n’existent que chez les anoures.
Après un temps variable (de une à quatre heures) et selon les espèces, on assiste à un mouvement de bascule de la calotte pigmentaire d’un angle d’environ 30°.
Il va ainsi libérer une région légèrement dépigmentée, le croissant gris, qui donnera la future région dorsale de l’embryon.
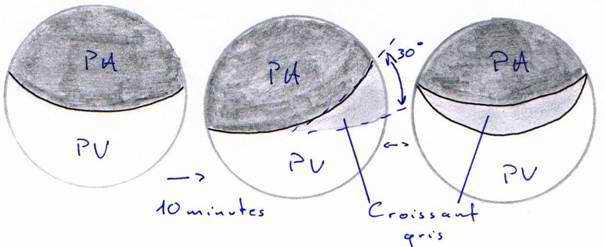
Remarque : chez les Urodèles, il y a un matériel visqueux entre la membrane plasmique et la paroi vitelline.
La région opposée au croissant gris donnera la région ventrale.
Le pôle animal et le pôle végétatif donnent respectivement la tête et la queue en formant l’axe antéropostérieur qui permettra la mise en place de l’axe dorso-ventral.
Cette rotation est appelée rotation de symétrisation et montre que l’embryon a délimité ses régions. Seule la région cytoplasmique tourne.
Lorsque la fécondation est monospermique, le croissant gris apparaît toujours diamétralement opposé au point de pénétration du spermatozoïde. Quand la fécondation est polyspermique, le croissant gris apparaît partout avec les mêmes potentialités.

La future face dorsale n’est pas une région très déterminée. Elle correspond à l’apparition d’une dissymétrie moléculaire qui entraîne une différence en un certain nombre d’acides nucléiques à cause de la rotation.
Le décollement de la membrane empêche la polyspermie. Le déplacement des molécules suppose l’initiation d’un mouvement et l’utilisation d’un support. Au cours de la rotation de symétrisation, on constate la formation d’un réseau de microtubules orientées parallèlement à la surface de l’œuf et servant de support au déplacement des déterminants moléculaires. Il y a aussi des molécules fournissant de l’énergie.
Tout ceci plaide en faveur d’une localisation moléculaire différentielle déterminant la future face dorsale de l’embryon. Du cytoplasme de la région du croissant gris qui est injecté sur la face ventrale d’un autre embryon va induire un deuxième embryon (siamois). Réciproquement, si l’on injecte du cytoplasme de la face ventrale, il ne se passe rien.
III La segmentation.
C’est une phase de divisions cellulaires : on passe d’un œuf unicellulaire à une blastula (6 à 10000 cellules). Cette segmentation implique de nombreux facteurs qui vont orienter les fuseaux.Le premier plan de division passe par l’axe pôle animal/pôle végétatif, dit méridien et donne 2 blastomères égaux. Ce plan peut couper, ou non, le croissant gris.
Le deuxième plan de clivage est parallèle au premier et donne 2 méridiens égaux.
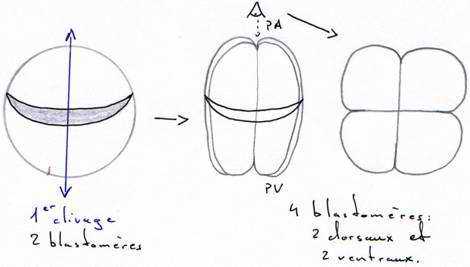
La segmentation est totale, et, pendant les deux premières divisions, elle est égale.
Au cours du troisième cycle cellulaire, la segmentation devient inégale (subéquatoriale) et donne 4 micromères au pôle animal et 4 macromères au pôle végétatif.
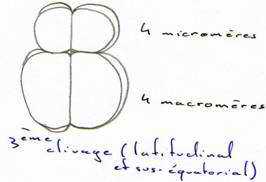
Les cycles cellulaires se poursuivent rapidement, sont synchrones jusqu’à la 10ème ou 12ème division puis, deviennent asynchrones. Ce cycle qui fait passer de la division synchrone à la division asynchrone est appelé transition blastuléenne.
Les cycles sont rapides jusqu’à la mi-blastula (pas de phase G1 ni G2).
Il y a une expression maternelle des gènes : toutes les protéines synthétisées le sont à partir d’ARN maternel stocké. Après la transition blastuléenne, les cycles s’allongent et apparaissent les phases G1 et G2 (G1, S, G2, M).
On a l’apparition d’un asynchronisme des divisions cellulaires. C’est une expression zygotique des gènes.
Au départ, on a un stade morula. Les cellules deviennent de plus en plus petites et vers la mi-segmentation, on aura le stade blastula.
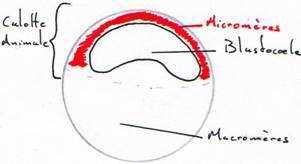
Il y a apparition d’une cavité de segmentation. Le stade blastula présente une cavité excentrée vers le pôle animal : c’est le blastocœle.
On a trois populations cellulaires qui se mettent en place pendant la gastrulation.
Cette blastula va synthétiser des molécules. A la fin de la segmentation, les micromères (du pôle animal) vont former la matrice extracellulaire qui permettra la suite des événements. Un des composants principaux est la fibronectine qui rendra possible la migration des cellules au cours de la gastrulation.
IV La gastrulation.
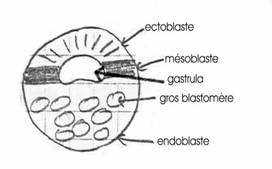
Il y a mise en place de trois feuillets :- ectoblaste
- mésoblaste
- endoblaste
A Les phénomènes externes de la gastrulation.
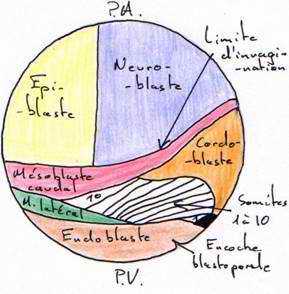
Il y a apparition d’une encoche blastoporale sous l’emplacement du croissant gris. Cette encoche résulte d’une invagination des cellules médiodorsales : c’est le stade grand bouchon vitellin.Au stade petit bouchon vitellin, le blastopore devient petit.
A la fin de la gastrulation, il ne persiste qu’une fente : l’anus.
B Les phénomènes internes de la gastrulation.
L’analyse des mouvements morphogénétiques de la gastrulation a été rendue possible par l’utilisation de marqueurs colorés (ex : VOGT) : c’est l’aboutissement à la carte des territoires présomptifs.- Les marques 1 à 4 : elles sont réparties dans la partie ventrale du système nerveux
- La marque 5 est dans le mésoderme caudal.
- La marque 6 est dans la corde dorsale.
- La marque 7 est dans le mésoderme précordal.
- Les marques 8 à 11 sont localisées dans la paroi ventrale du tube digestif.
Le mésoblaste donnera la musculature, la corde et des feuillets qui participent à l’emballage.
C Les mouvements morphogénétiques de la gastrulation.
A la partie inférieure médiane du croissant gris, un mouvement d’invagination se constitue, provoquant l’apparition d’une nouvelle cavité : l’archantéron.Le blastocœle disparaît à la fin de la gastrulation.
Cette invagination intéresse un peu d’endoblaste, la plaque précordale (pharynx) puis le cordomésoblaste ainsi que le mésoblaste somitique et latéral. En fin de gastrulation, le blastocoele a complètement disparu et il est remplacé par l’archantéron qui est la future lumière du tube digestif.
Quand la gastrulation est terminée, les lames mésodermiques droite et gauche ne se sont pas encore rejointes. Elles ne le feront qu’au cours de la neuralation.
D’un germe à deux ensembles cellulaires (pôle animal et pôle végétatif), on passe progressivement, au cours de la gastrulation, à trois ensembles cellulaires qui s’emboîtent les uns dans les autres pour donner trois feuillets fondamentaux. En raison de la position respective de ces trois feuillets, on parlera d’ectoblaste à l’extérieur, de mésoblaste en position intermédiaire et d’endoblaste à l’intérieur.
On a différents types de mouvements :
- épibolie à allongement
- de convergence
- divergence des lames latérales
- d’épibolie de l’ectoblaste pour compenser l’invagination du mésoblaste et de l’endoblaste
- d’extension au niveau du mésoderme somitique.
L’embryon, à la fin de la gastrulation, a toujours une forme cylindrique et le blastopore constituera toujours l’anus.
V La neurulation (mise en place de la plaque neurale).
Le processus d’induction neurale se fait lors de la gastrulation. Le cordomésoblaste vient au contact de l’ectoderme et induit celui-ci en plaque neurale (neurectoderme). Le cordomésoblaste va libérer des molécules qui permettent la différenciation et l’activation des gènes.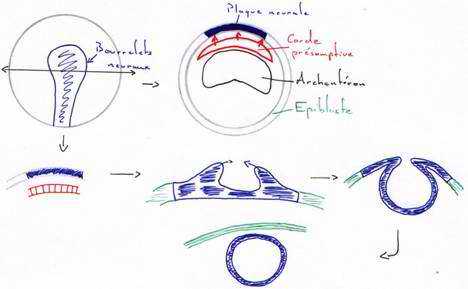
L’ectoblaste est compétent pour capter les signaux (moléculaires), mais cette compétence est transitoire.
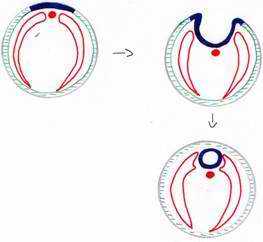
Au début de la neurulation, il y a mise en place de la plaque neurale. On assiste en premier à des mouvements pigmentaires sur la face dorsale qui dessinent une aire en forme de raquette.
Ces traînées pigmentaires sont dues à un épaississement du neurectoderme en plaques neurales. Des bourrelets neuraux vont s’épaissir, se soulever puis se souder pour former le tube neural. La fermeture se fait d’abord dans la région du tronc puis dans celle de la queue (région caudale) et enfin, dans la région antérieure. Ce tube nerveux donnera l’encéphale et la moelle épinière. Ces cellules des crêtes neurales ont des capacités de migration importante grâce à la synthèse de matrice extracellulaire sur laquelle elles migrent.
Ces cellules qui vont migrer vont diffuser dans tout le corps et donner, des mélanophores (cellules sécrétant des pigments) qui donneront sa couleur a l’embryon, les ganglions spinaux (ou rachidiens), les systèmes sympathiques, les glandes endocrines (médullo-surrénales).
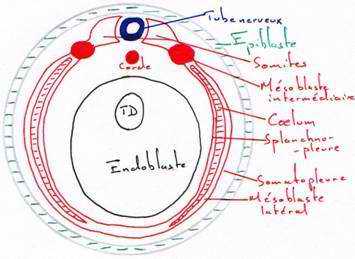
Pendant que se déroule la neurulation, on assiste à une régionalisation du mésoblaste en lame cellulaire pleine formant la voûte et les parois latérales de l’archantéron. Alors que les lames latérales se rejoignent ventralement, le mésoderme dorsal s’individualise, la corde s’isole et le mésoderme dorsal se métamérise en somites (blocs métamérisés). La région dorsale externe va donner le dermatome (derme de la peau). La région supérieure interne va donner le myotome (muscle strié), la partie inférieure interne donnera le sclérotome (les vertèbres). La région intermédiaire donnera les gonades et les reins.
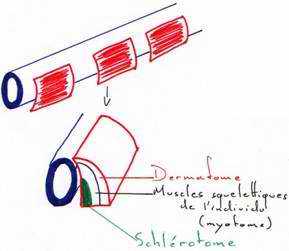
Les régions latérales et la région ventrale du mésoderme ne se métamérisent pas et se délaminent en deux feuillets :
- le plus externe, le somatopleure, donnera le squelette des membres, le péricarde et les muscles viscéraux
- la splanchnopleure va donner le cœur, les premières cellules sanguines, la paroi endothéliale des vaisseaux sanguins.
Ces deux feuillets vont constituer une cavité cœlomique (cavité générale du cœur).
VI La formation du tube digestif.
A la fin de la gastrulation, la masse d’endoblaste constitue le plancher de l’archantéron.Pendant la neurulation, les bords latéraux de l’endoblaste se soulèvent et viennent se souder dorsalement dans un mouvement inverse à celui du mésoblaste latéral.
L’endoblaste donnera l’épithélium du tube digestif, de l’appareil urinaire et pulmonaire. Celui-ci est étroite association avec le mésoderme splanchnique. L’endoblaste donnera des glandes annexes comme le foie, le pancréas.
Au cours de la neurulation, le germe s’allonge dans le sens antéropostérieur. A la fin de cette neurulation, on distingue la tête et la queue : c’est le stade du bourgeon caudal. Divers organes apparaissent dans la région antérieure comme les yeux, les branchies, les ventouses adhésives pour les anoures ou le balancier pour les urodèles.
On assiste à une régionalisation du système nerveux donnant l’encéphale et la moelle épinière. Cet encéphale va devenir inducteur vis à vis de l’ectoblaste. C’est ainsi que le proencéphale induit la placode (vésicules) olfactive.
Le diencéphale va former 2 invaginations latérales constituant la rétine qui induit la formation du cristallin qui induira, lui-même, la formation de la cornée (cascade d’inductions).
Le rhombencéphale induit la placode auditive ou otique. Les tissus et les organes se mettent tous en place.