
I Les phytohormones.
 Les phytohormones sont des substances chimiques synthétisées par le végétale. Elles sont oligodynamiques : actives à faible dose. Elles agissent parfois à distance du lieu de synthèse : elles ont une influence sur le fonctionnement.
Les phytohormones sont des substances chimiques synthétisées par le végétale. Elles sont oligodynamiques : actives à faible dose. Elles agissent parfois à distance du lieu de synthèse : elles ont une influence sur le fonctionnement.A Les auxines.
1 Nature et répartition dans la plante.
Les auxines sont synthétisées dans l’apex des tiges et des jeunes feuilles. Elles sont libres ou liées à d’autres molécules. Leur transport par le phloëme est polarisé : c’est un transport actif basipète (du haut vers le bas). Ces molécules sont des composés instables, sensibles à la lumière, à l’acidité, à l’oxygène.
2 Mode d’intervention des phytohormones.
Les effets des phytohormones sont moins prononcés chez les végétaux que chez les animaux (demande de réponse précise). Chez les végétaux, la transmission des informations est moins précise. On a du mal à séparer la transduction du signal et sa conséquence.
Les récepteurs aux phytohormones sont localisés, soit dans le cytoplasme (et accompagnent les hormones jusqu’au lieu d’action), soit sur/dans les membranes et ont alors un rôle de messagers secondaires.
L’Inositol-triphosphate : IP3 et le complexe Ca2+/calmoduline.

Au niveau de la membrane, les phospholipides sont transformés en PiP2. L’hormone reconnaît son récepteur et active dans la membrane, une enzyme (une lipase), qui hydrolyse le PiP2 en IP3 et DiAcylGlycérol. L’IP3 permet l’ouverture de canaux calciques au niveau de la vacuole et du réticulum plasmique : on a alors formation du complexe Ca2+/calmoduline qui va activer une kinase qui ensuite, va phosphoryler un composé «E». Ce composé « E » va assurer le transfert de l’AIA (hormone) vers le noyau.
3 Propriétés physiologiques.
Les effets sur l’élongation sont différents si l’on se trouve sur une tige ou une racine. Une dose favorable pour l’augmentation d’une tige peut-être défavorable pour l’augmentation d’une racine. En règle générale, les auxines favorisent l’accroissement des racines alors que les cytokinines favorise la croissance des tiges et des bourgeons. L’élongation fait appel à la plasticité de la paroi et à sa structure fibrillaire dans laquelle s’entrecroisent des chaînes de molécules diverses, où, entre ces chaînes, se crées des liaisons. Pour qu’il y ait élongation, il faut couper les liaisons interchaînes (intervention de l’auxine) au niveau périphérique et nucléaire.
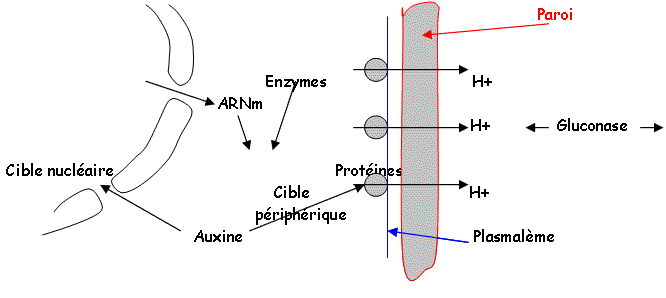
Le complexe récepteur de l’AIA dans le plasmalème, va stimuler les ATPases de celui-ci. On assiste à l’expulsion de protons vers la paroi ainsi qu’à une diminution du pH dont le but est la coupure des liaisons hydrogène.
La gluconase va couper les chaînes de polysaccharides. L’auxine va activer les pompes à protons, ce qui entraîne la possibilité de l’extension de la paroi.
La cible nucléaire : l’AIA et le récepteur soluble activent dans le noyau les gènes des enzymes responsables de la synthèse des matériaux comme les polysaccharides (pariétaux). On assiste alors à l’extension de la paroi.
En utilisant les inhibiteurs, on s’aperçoit que l’action de l’auxine se fait au niveau de la transcription.
B Les gibberellines.
1 Nature et répartition dans la plante.
Les gibberellines sont des diterpènes (molécules à 20 carbones), formées à partir de l’isoprène. Elles ont été découvertes en étudiant la maladie du ris (maladie due à un champignon).
La synthèse de ces molécules est effectuée dans des sites divers mais toujours dans des lieux de division active (à l’apex de tiges et de racines). Leur migration est polarisée. Elle se déroule dans le xylème et dans le phloëme, toujours liée à des sucres.
2 Propriétés physiologiques.
a Effets sur la croissance.
Les gibberellines permettent un allongement des entre-nœuds. Leur action se fait en synergie avec les auxines et les cytokinines.
b Autres effets.
Elles permettent la levée de la dormance des semences. Elles ont un effet antagoniste à celui de l’ABA.
c Applications.
On a trouvé des anti-gibberellines, qui sont des nanifiants : CCC, Amo16-18, phosphon : ils bloquent la synthèse des gibberellines. Les plantes ainsi traitées ont un aspect nain (au niveau des tiges mais jamais au niveau des feuilles).
C Les cytokinines.
1 Nature et répartition.
Ce sont des substances qui augmentent la division cellulaire. La première cytokinine découverte fut la zéatine, isolée dans le mais.
Ces hormones sont trouvées dans tous les tissus et sont synthétisées sur les lieux d’utilisation. On observe de fortes synthèses au niveau des racines.
2 Propriétés physiologiques.
a Au niveau cellulaire.
Les cytokinines sont présentes dans la sève brute et transportées sous forme de nucléotides de zéatine. Elles stimulent la division cellulaire en présence d’auxine : action en synergie.
b Au niveau de l’organisme.
On les trouve :
- au niveau des bourgeons, elles sont antagonistes des auxines
- pour la levée de dormance des graines,
- au moment du développement des ébauches florales,
- dans les feuilles, où elles favorisent la diminution du turn-over de la chlorophylle, en améliorant le verdissement des feuilles.
D L’acide abscissique.
1 Nature et répartition.
L’acide abscissique est un composé en C15 qui a été trouvé pendant l’abscission des fruits et des feuilles de cotonnier. Sa synthèse est réalisée au niveau des graines et des semences.
2 Propriétés physiologiques.
Il est antagoniste des gibberellines, en prolongeant la dormance des graines et des bourgeons. Il intervient aussi dans la fermeture des stomates. Il favorise la maturation des graines en stimulant l’accroissement de l’embryon et l’augmentation des réserves tout en inhibant la germination.
E L’éthylène.
On dose ce qui est dégagé par la plante. Il se forme à partir de méthionine. Il est antagoniste aux gibberellines et à un rôle dans le mûrissement des fruits.
Il inhibe l’élongation en diminuant les tiges et les racines : c’est une hormone de stress, de défense.
II Le phytochrome.
A Nature.
1 Mise en évidence
Chez les végétaux, on trouve des molécules différentes de la chlorophylle, qui sont capables de capter la lumière et d’induire sa transduction pendant le développement des végétaux. L’accroissement de la plante est modelé par la lumière : c’est la photomorphogenèse (ensemble des réactions végétales réalisées grâce à la lumière).
S’il y a absorption d’une lumière monochromatique rouge à faible énergie : le rouge clair déclenche une réponse physiologique qui est annulée par le proche infrarouge (rouge sombre). C’est la qualité du dernier éclairement qui prédomine.
Il existe dans la plante, un pigment qui, à l’obscurité, est capable d’absorber la lumière rouge. Ce pigment se transforme en une forme qui, à son tour, absorbera le rouge sombre.
La forme capable d’absorber le rouge sombre déclenche la réponse physiologique (croissance des tiges et des feuilles et arrêt de la floraison). Ces formes de pigments sont photo-convertibles.
2 Constitution.
Chromophore + protéines → chaîne tétrapyrolique → absorption de lumière par modifications de la position de certains hydrogènes et doubles liaisons.
Ce phénomène est comparable à ce qui se passe dans l’œil. La lumière blanche a le même effet que la lumière rouge en réalisant une transformation qui est relativement lente (1/10ème de seconde).
B Répartition dans la plante.
Les phytochromes sont présents en faible quantité dans le végétal. Le jour, ils sont sous forme P730 (ils absorbent à λ=730nm). La nuit, ces phytochromes sont sous leur forme P660.
La forme active est la forme P730. Pendant la nuit, elle se convertie en forme inactive en subissant un turn-over. Ce pigment est répandu chez les végétaux supérieurs (phanérogames, bryophytes et ptérydophytes).
C Les modalités d’action.
Il semble que le phytochrome agisse sur les modalités cellulaires par un changement de configuration qui modifie les propriétés membranaires, rendant possibles le passage de métabolites, de cations, d’hormones et d’ATP.
Au niveau nucléaire, les phytochrome agissent sous l’activation ou la répression de certains gènes. Quand ils sont sous forme active, ils permettent l’activation de kinases.
D Phénomènes contrôlés par le phytochrome.
La photomorphose positive (>0) ; exemple : activation par P730.
La photomorphose négative (<0 :="" exemple="" inhibition="" p730.="" p="" par="">
La photomorphose mixte ; exemple : le photopériodisme.
La photomorphose positive : pour les plantes qui germent à la lumière, le contrôle de la germination est réalisée par le phytochrome.
L’accroissement des feuilles est stimulé par la lumière rouge et inhibée par le rouge sombre.
L’action sur le mouvement des feuilles : de nombreuses feuilles pennées (chez les légumineuses) subissent des mouvements nyctinastiques (c’est le mouvement des feuilles qui se plient). Ce mouvement a lieu si la quantité de forme active est suffisante en fin de journée. Le mouvement est due à des variations différentielles de turgescence (cellule à la base des folioles).
La synthèse des pigments entraîne la synthèse d’enzymes : PAL (PhénylalanineAmmoniaLyse), PEPcarboxylase, Rubisco.
La photomorphose négative : c’est l’inhibition de la croissance des tiges. Il y a synthèse de Lipoxygénase qui va permettre la production d’éthylène. La photomorphose mixte : elle déclenche une induction ou une répression. L’exemple type en est la floraison.