

Chez les métazoaires triploblastiques, il y a apparition du mésoderme (troisième feuillet embryonnaire) qui dérive de l’endoderme pendant le développement embryonnaire. L’ectoderme et l’endoderme ont des fonctions bien définies ; le mésoderme permet la différenciation d’organes internes (appareil excréteur, organes génitaux) dont la musculature qui permettra des mouvements orientés (locomotion). Il apparaît le déplacement dans une direction donnée : les animaux ont maintenant un avant et un arrière.
Apparition de la symétrie bilatérale.
La symétrie bilatérale remplace la symétrie radiaire des cnidaires ; l’animal a donc une région antérieure, une postérieure et une orientation dorso-ventrale. Cette symétrie existe à l’état embryonnaire mais peut disparaître chez l’adulte comme chez les oursins (pentaradiés) ou chez les gastéropodes.Cette symétrie bilatérale est un des principaux facteurs qui vont infléchir le cours de l’évolution animale.
Le déplacement se fait tête vers l’avant.
La céphalisation.
La céphalisation est un développement des appareils sensoriels et du système nerveux : elle favorise la coordination des mouvements. Les organes sensoriels se regroupent dans la région antérieure, au voisinage d’un système nerveux principal (encéphale). C’est le développement de la tête s’il y a un « cerveau », des organes sensoriels et d’une bouche.Chez les triploblastiques, il existe deux groupes : les acœlomates et les cœlomates. Ces groupes sont différenciés par la structure du mésoderme:
- Si le mésoderme donne un tissu épais, on se trouve chez les acœlomates.
- Si le mésoderme donne un tissu creux favorisant l’apparition de vésicules cœlomiques, on se trouve chez les cœlomates.
I Les Acœlomates.
Le mésoderme vient remplir les espaces entre les organes : c’est un parenchyme.A Les Plathelminthes.
Les plathelminthes sont les triploblastiques les plus primitifs. Ils présentent un aplatissement dorso-ventral avec, évidemment, l’existence des faces dorsale et ventrale. Le corps est organisé en deux territoires différents :- La région céphalique, antérieure (fonctions sensorielles).
- La région postérieure, caudale.
L’appareil respiratoire est absent : les échanges gazeux s’effectuent à travers le tégument de l’animal, ou bien, comme chez les parasites, il n’y aura pas d’échange gazeux.
L’appareil digestif est soit incomplet, soit absent. S’il est incomplet, c’est qu’il lui manque l’anus : « appareil en cul de sac ». S’il est absent, les échanges seront possibles par osmose (pour les parasites).
Les plathelminthes comprennent environ 50 000 espèces et peuvent être libres ou parasites. Il existe six classes mais nous n’en verrons que 4 ici : Turbellariés (planaires), Trématodes (douves), Cestodes (ténia) et Monogènes (polystomiens).
1 Les Turbellariés.
Les turbellariés sont essentiellement aquatiques (eau douce / marin) avec quelques formes terrestres (dans les forêts tropicales humides). Leur taille est souvent inférieure à 2 centimètres et peuvent exceptionnellement atteindre 60 millimètres.Ils sont libres et non segmentés. Ils présentent une région antérieure avec deux yeux (ou plus) et des auricules. Ils ont une région spécialisée au moment de la reproduction.
α Le tégument.

Entre les couches musculaires, on distingue des cellules glandulaires qui peuvent émettre des digitations vers l’épithélium. Au niveau de la membrane basale, on a localement des cellules sensorielles.
Les cellules épithéliales ont un petit corpuscule (rhabdite) à rôle encore inconnu. Elles produisent un mucus à l’extérieur de l’animal pour enrober les corps étrangers.
Sous les couches musculaires, on a deux types cellulaires :
- Des cellules amiboïdes qui permettent les échanges respiratoires et nutritionnels.
- Des cellules étoilées qui baignent dans l’hémolymphe.
ß Système nerveux et organes sensoriels.

Des nerfs sensitifs partent aussi en direction des yeux et des auricules.
Au niveau des yeux est présent un tégument transparent qui permet l’entrée de la lumière. L’œil est composé d’une cupule tapissée de cellules pigmentées. Cette cupule concentre la lumière vers les cellules rétiniennes. L’œil a une disposition inversée.
Les auricules ont un rôle tactile et parfois de préhension.
Il existe cinq récepteurs :
- Les tangorécepteurs: ils ont un rôle tactile lors des contacts avec des surfaces solides.
- Les rhéorécepteurs : ce sont des cellules sensorielles appréciant les courants.
- Les chémorécepteurs : ils détectent les composés chimiques.
- Les statorécepteurs: ils ont un rôle dans l’équilibration.
- Les photorécepteurs: ils sont sensibles à la lumière.
Exemple de statorécepteurs : les statocystes. Ils ont la forme d’une vésicule avec un statolithe et autour, une cellule spécialisée : un lithocyste.
γ L’appareil digestif.
")
Il existe trois types de pharynx :
- Le pharynx simple : il est formé par une invagination de l’ectoderme (épithélium cilié) avec une fine paroi musculaire.
- Le pharynx bulbeux: il est toujours avec un épithélium cilié mais la musculature est plus développée que précédemment (pour la mastication).
- Le pharynx plissé: il est en forme de trompe dévaginable et rétractable, ce qui permet la capture de proies ou la succion de grosses proies.
δ L’appareil excréteur

A l’extrémité de toutes les canalicules, on trouve une cellule excrétrice rudimentaire : protonéphridie, d’origine ectodermique. Ce type de cellule est aussi appelé « cellules à flamme ». Elles sont ciliées et c’est le battement de leurs cils qui pousse les déchets venant de l’hémolymphe dans des canaux collecteurs en direction des néphridiopores.
Cet appareil excréteur sert aussi à sortir l’eau du corps pour maintenir l’équilibre osmotique. Tout cela est vrai pour les formes d’eau douce qui ont un grand besoin de régulation osmotique. Ce n’est pas vrai pour les formes marines.
Le nombre de néphridiopores est différent selon le groupe de turbellariés (1, 2 ou plus).
ε L’appareil reproducteur et le développement.
Les turbellariés sont hermaphrodites.L’appareil sexuel mâle est composé de testicules fragmentés en follicules testiculaires drainés par deux spermiductes (ou canaux déférents) qui s’élargissent pour former les vésicules séminales à fonction de stockage des produits génitaux.
Ces deux vésicules confluent pour former un pénis tubulaire et musculeux. Ce dernier débouche dans l’atrium génital, commun aux deux sexes.
L’appareil sexuel femelle est constitué par deux ovaires en position antérieure, drainés par des oviductes (ou vitelloductes) où l’on trouve de nombreuses glandes vitellogènes qui produisent les cellules vitellines, cellules nourricières pour l’œuf.
Lors de la reproduction, les gamètes mâles et femelles confluent vers l’atrium génital où il existe un réceptacle séminal (ou « bourse copulatrice ») qui favorise le mélange des gamètes. Toutefois, il n’y a pas d’autofécondation car ce sont des hermaphrodites protandres (la maturité sexuelle mâle arrive avant la maturité sexuelle femelle).
Les œufs se retrouvent dans des cocons, par groupe de 4 à 6 où l’on trouve de nombreuses cellules vitellines.
La segmentation de l’embryon est de type spirale. Le développement est direct, sans stade intermédiaire larvaire.
ζ La multiplication asexuée.
La multiplication asexuée peut se faire selon deux modalités :- Par architomie : il y a division d’un individu en deux puis régénération des parties manquantes.
- Par paratomie : il y a d’abord régénération puis division.
η Systématique.
La systématique est basée sur la forme et sur la structure de l’appareil digestif. On distingue :- L’ordre des Acèles (ou Acœles) : l’appareil digestif est réduit. On y trouve la bouche ainsi qu’une petite cavité buccale (il n’y a ni pharynx, ni intestin). Les formes de cet ordre sont essentiellement marines.
- L’ordre des Rhabdocèles : ils ont une bouche, un pharynx et un intestin rectiligne simple.
- L’ordre des Polyclades : L’intestin est très développé et ramifié. Les ramifications sont rayonnantes par rapport au centre de l’animal.
- L’ordre des Triclades: L’intestin est formé de
trois branches (une antérieure et deux postérieures). De plus, chacune
des branches est diverticulée. Chez les triclades, on trouve les
planaires au sens strict. On pourra distinguer trois sous-ordres en
fonction de l’écologie des espèces:
- Les Terricoles sont terrestres.
- Les Maricoles sont marins.
- Les Paludicoles sont d’eau douce.
2 Les Trématodes.

- Il y a perte de la ciliature de l’épithélium (pas de déplacement).
- Il y a régression des organes des sens (en relation avec la perte de la locomotion) : on va parler d’une « évolution régressive » observée chez les Trématodes.
- Il y a développement d’organes de fixation qui sont souvent une ventouse buccale antérieure et une ventouse ventrale ou « acétabulum ».
Le développement est indirect avec de nombreuses formes larvaires asexuées : il y a un développement emboîté chez les trématodes.
a Le tégument.

La présence d’une cuticule a deux principales conséquences : le développement ciliaire est impossible sur l’épithélium ; la croissance est alors discontinue et est réalisée par mues successives.
La cuticule a une lame basale. En dessous, on trouve un parenchyme et différents faisceaux musculaires (circulaires et longitudinaux). Sous cet ensemble, on trouve des cellules épidermiques qui peuvent émettre des diverticules jusqu’à la cuticule. Les cellules épidermiques n’ont pas de membrane cellulaire : c’est un tégument syncitial.
ß Système nerveux.
Le système nerveux est concentré en arrière de la ventouse buccale. Sa structure est proche de celle des turbellariés, avec des ganglions cérébroïdes reliés par une commissure. Des terminaisons nerveuses partent vers l’avant et des cordons nerveux partent vers l’arrière.Différence : il y a absence d’organe des sens car les trématodes sont des parasites. Il leur reste quelques cellules sensorielles spécialisées.
&gama; L’appareil digestif.
La bouche est antérieure, terminale ou sub-terminal et s’ouvre au centre de la ventouse buccale.L’intestin est formé par deux branches (cæcum digestif), très ramifiées où vient s’accumuler le sang ingéré.
Il n’y a pas d’anus.
δ L’appareil excréteur.
La structure de l’appareil excréteur est identique à celle des Turbellariés (protonéphridien) avec un réseau caniculaire dense. Les canicules se réunissent en deux troncs principaux qui débouchent dans un réservoir terminal (« vessie ») qui s’ouvre à l’extérieur, par l’orifice excréteur (en position caudale).ε L’appareil reproducteur.
Les Trématodes sont hermaphrodites.- L’appareil mâle : Cet appareil est formé de deux testicules (ou follicules testiculaires) drainés par deux spermiductes qui convergent en une vésicule séminale de stockage. De cette vésicule se différencie un pénis copulateur qui fait saillie au niveau du gonophore (orifice mâle et femelle).
- L’appareil femelle : L’appareil femelle est relativement complexe. Les fonctions vitellogènes et gamétogènes sont dissociées dans l’espace. Il gravite autour d’une chambre (l’ootype) où arrive l’oviducte, venant d’un ovaire ramifié. L’ootype reçoit un canal issu de la fusion des deux vitelloductes qui amènent les cellules vitellines venant des glandes vitellogènes (très ramifiées).
La fécondation se fait par accolement réciproque.
L’œuf fécondé est entouré d’une coque et accompagné de nombreuses cellules vitellines. La reproduction s’effectue au niveau de l’hôte intermédiaire (pendant le cycle). Un œuf donnera alors de nombreux individus.
η Le cycle parasitaire.
Ce cycle comporte de nombreux stades : on parle alors de « développement emboîté ». Les diverses formes larvaires viennent du cycle théorique des Trématodes (plusieurs stades larvaires avant le stade adulte).
- donner des cercaires, ce qui est le 4ème stade larvaire,
- ou donner des rédies filles, ce qui permet une multiplication du nombre de rédies.
A ce stade, il y a sortie de l’hôte intermédiaire puis enkystement qui donne la forme métacercaire : c’est le stade infectant de l’hôte définitif, qui donnera l’adulte.
Un œuf donnera de nombreux individus grâce aux diverses multiplications larvaires asexuées du cycle.
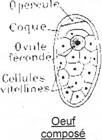
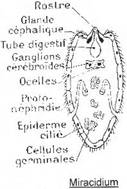
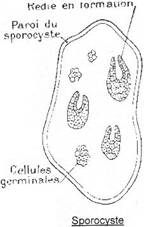
Exemple : la petite douve du foie, Dicrocœlium dendriticum.
Cette douve vit dans les canaux biliaires du mouton. L’adulte mesure environ 10 millimètres de long et 2 à 3 millimètres de large. Il existe deux hôtes intermédiaires :
- Un gastéropode terrestre (Hélix)
- et une fourmi (Formica).
La petite douve peut infecter l’Homme par ingestion de foie de mouton mal cuit, mais c’est une infestation accidentelle.
Exemple : La grande Douve du foie, Fasciola hepatica.
L’adulte se trouve dans le foie de mouton. L’hôte intermédiaire est un gastéropode aquatique (Limnée) infesté par le stade miracidium (qui est libre et nageur dans le milieu aquatique).
La larve miracidium, par enkystement en sporocyste, donne des rédies. Selon les conditions de température, les rédies ont deux devenirs possibles : si l’eau a une température supérieure à 25°C, elles redonnent des rédies (rédies filles) ; si la température est inférieure à 25°C, on passe au stade cercaire.
Le stade cercaire se fixe et s’enkyste en métacercaire sur les plantes aquatiques comme le cresson. Si l’Homme consomme des végétaux souillés, il peut être contaminé et être atteint de distomatose. Toutefois, l’Homme n’est pas indispensable au cycle de Fasciola.
Exemple : la douve pulmonaire.
Cette douve provoque des bronchites sanglantes ressemblant à la tuberculose ou à la pneumonie. Son cycle fait intervenir : l’Homme, un mollusque (Melania) et un crabe (Potamon).
Le mollusque est le lieu de développement de stades larvaires, jusqu’au stade cercaire. Le crabe permet le passage du stade cercaire au stade métacercaire.
Exemple : Schisostome (bilharziose).

L’hôte intermédiaire est un mollusque où la larve miracidium donne un sporocyste puis le stade cercaire (nageur) qui sera libéré. Les furocercaires vont contaminer l’Homme en pénétrant par la peau lors d’une baignade (15 minutes sont nécessaires pour qu’un furocercaire rentre complètement).
Dans ce cycle, il y a absence des stades rédies et métacercaires : c’est une diminution des formes larvaires. Le cycle devient plus rapide avec moins d’évènements hasardeux.
3 Les Monogènes (= Polystomiens).

a Morphologie.
La bouche est antérieure ou ventrale. Il y a plusieurs ventouses : la ventouse antérieure est appelée « cotylophore » ; les autres ventouses sont postérieures, accompagnées de crochets.On note la présence d’un canal génito-intestinal servant à évacuer le trop plein de cellules vitellines.
b Exemple : Polystoma integerrinum.
C’est un parasite de la grenouille rousse (Rana temporaria). Il y a un synchronisme entre les cycles biologiques de ces deux espèces (certaines phases du cycle sont conjointes). Au moment de la reproduction (ponte) de la grenouille, les Polystoma adultes pondent des œufs dans les urines (et sont évacués en milieu aquatique).Ces œufs donnent une larve ciliée nageuse ; un oncomiracidium qui peut se fixer sur les têtards de grenouille rousse.
Si le têtard a plus de 10 jours, la larve gagne l’appareil digestif du têtard et va atteindre la vessie pour donner un adulte. L’adulte est apte à se reproduire au bout de trois ans, comme les batraciens.
Si la larve rencontre un têtard de moins de 7 à 10 jours, la suite du cycle parasitaire ne peut avoir lieu : pas d’infestation.

c Exemple: Diplozoon paradoxum.

Les larves sont aspirées par les poissons et se fixent sur les branchies. Elles y subissent une métamorphose qui différencie une ventouse circulaire dans la partie médiane ventrale (de la larve) et un petit bouton sur la face dorsale, formé par un soulèvement de la cuticule. Ce sont des larves diporpas qui se nourrissent du sang du poisson pendant plusieurs semaines. La suite du développement se fait si deux larves entrent en contact l’une avec l’autre. L’accolement se fait par la ventouse qui attrape le bouton, chez l’un comme chez l’autre.
Si l’accolement se fait, il y a fusion de ces larves au niveau de leurs points d’attache : c’est une anastomose entre les deux intestins ; la nourriture ingérée par l’une est alors utilisable par l’autre. Il y a aussi fusion des conduits sexuels mâles et femelles.
Les deux individus sont définitivement accouplés. La fusion met en connexion les canaux déférents de l’un avec le vagin de l’autre : c’est une fécondation réciproque croisée.
4 Les Cestodes (classe).
Les cestodes sont des parasites du tube digestif de vertébrés supérieurs. Leur corps est aplati, segmenté, plus ou moins rubané. Ce sont des animaux généralement longs : 12 à 20 centimètres pour les plus grands. Ils n’ont ni tube digestif ni appareil respiratoire.a Morphologie.
Le corps est divisé en trois parties :- Région antérieure. C’est le scolex ; il porte le dispositif de fixation : ventouse et/ou crochets en couronne.
- Zone de prolifération ou cou. C’est à ce niveau que se forment les segments composant le corps de l’animal.
- Le strobile. Il occupe 9/10ème du corps. Il est formé d’une succession de segments (les proglottis).
b Le tégument.

c Le système nerveux.
Le système nerveux est situé dans le scolex et est constitué de deux ganglions cérébroïdes présentant des prolongements antérieurs et postérieurs.Les prolongements antérieurs s’unissent par une commissure circulaire où se situent deux ganglions dorsaux et deux ventraux.
De chaque ganglion part un cordon nerveux, à l’intérieur du strobile.
Vers l’arrière, on trouve une commissure avec deux ganglions latéraux d’où partent deux cordons latéraux dans le strobile.
A l’extrémité postérieure des proglottis, on a une liaison (commissure) qui relie les six cordons nerveux avec les deux ganglions.
Il n’y a pas d’organe sensoriel différencié mais de nombreuses terminaisons nerveuses aboutissent à la cuticule.
d L’appareil excréteur.
Cet appareil excréteur est composé de 4 canaux excréteurs tout le long du strobile : deux ventraux et deux latéraux.Dans la région postérieure de chaque proglottis, on a une anastomose qui réunie les canaux ventraux. Ils débouchent à l’extérieur, au niveau du dernier proglottis du strobile alors que les canaux dorsaux s’atrophient avant d’atteindre l’extrémité du strobile.
e L’appareil reproducteur.
L’appareil reproducteur rappelle celui des Trématodes ; il y a hermaphrodisme. Un pore génital est commun aux deux sexes : c’est l’atrium génital. L’hermaphrodisme est protandre. On trouve les appareils mâle et femelle dans le même proglottis.Les proglottis antérieurs porteront l’appareil mâle.
Les proglottis moyens auront un appareil mâle et un appareil femelle bien développé.
Les proglottis âgés auront un appareil mâle dégénéré et un développement important de l’utérus qui se ramifie et va contenir de nombreux œufs en développement (5000 œufs par utérus).
f Classification.
La classification des cestodes est réalisée à partir de la structure du scolex, de l’appareil génital et sur les particularités du cycle parasitaire. On distingue deux sous-classes :- Les Cestodaires : ils n’ont pas de scolex.
- Les Eucestodes: ils possèdent un scolex. Cette sous-classe regroupe deux ordres :
- L’ordre des Cyclophyllidiens. Le scolex possède quatre ventouses avec des fibres musculaires radiaires.
- L’ordre des Pseudophyllidiens. Le scolex présente deux dépressions (pseudobothridies), une dorsale et une ventrale. La musculature est peu différenciée.
g Développement.
Les cestodes sont dans le tube digestif de leur hôte définitif et s’y nourrissent par osmose et n’ont donc pas de tube digestif : ce sont des « osmotrophes ». Ils n’ont pas d’appareil respiratoire car ils ont une vie en anaérobiose.Pour pouvoir réaliser tout leur cycle de développement, ils se sont glissés dans la chaîne alimentaire. Ils ont un grand pouvoir reproducteur ; ils sont composés d’un grand nombre de segments (proglottis) formés par strobilation, ce qui est assimilable à une reproduction asexuée (fournie par des entités reproductrices complètes).

La fécondation est réalisée quand un repli de l’animal met en relation des proglottis de sexe différent. A la suite de la fécondation, les cucurbitains donnent des millions d’œufs. Toutefois, tous les cestodes ne sont pas aussi féconds : de petites espèces compensent leur faible nombre d’œufs par une multiplication asexuée à l’état larvaire.
Un œuf fécondé donnera par divisions, un embryon entouré de 3 ou 4 blastomères. Le tout est entouré d’une coque. L’embryon développe trois paires de crochets : c’est la larve hexacanthe enveloppée par une coque (embryonophore). Embryonophore + embryon hexacanthe = oncosphère. Cette dernière peut être aquatique et libre et elle sera appelée larve coracidium.
Le cycle de développement ne se poursuit que si le stade oncosphère est ingéré. Les sucs digestifs détruisent la coque, cassent l’embryonophore et libèrent l’embryon. Ce dernier se fixe par les crochets, perfore l’épithélium et se dirige vers la musculature par la voie sanguine. Une fois dans les muscles, il y a perte des crochets. Il se forme alors une invagination qui donnera le futur scolex : c’est un cysticerque.
Le stade cysticerque est infestant. S’il rencontre un deuxième hôte, il se transforme en procercoïde puis en plérocercoïde.
Le cysticerque est une vésicule sphérique, creuse, avec un scolex invaginé. Pendant sa phase d’attente, le cysticerque s’enkyste dans les muscles.
La larve procercoïde est une larve allongée, dépourvue de cavité interne. Son corps présente une constriction délimitant deux parties : à l’avant, on a la future larve et la partie postérieure dégénère.
La larve plérocercoïde possède le scolex du vers adulte et un débutde segmentation. Le corps est allongé (plusieurs millimètres de long), d’une couleur blanchâtre qui est due à la présence de particules calcaires dans le parenchyme.
h Cycles.
Tænia saginata.

Tænia solium.

Bothriocéphale : Diphyllobothrium latum.
Ce vers peut atteindre 2 à 8 mètres et même parfois, 20 mètres. Le scolex est ovoïde et porte deux pseudobothridies (c’est un pseudophyllidé). On y trouve entre 3000 et 4000 anneaux avec un pore génital en position médiane. Les adultes se trouvent dans l’intestin de l’Homme, du chat, du chien, du renard. Les oncosphères sont éliminées avec les selles. La larve est ciliée, nageuse à larve coracidium. Ce stade larvaire va être avalé par un crustacé du genre copépode dans l’intestin duquel il perd sa ciliature et puis, pénètre dans la cavité générale. C’est alors une larve procercoïde. Le développement s’arrête et la larve vit aussi longtemps que le crustacé. Si ce dernier est capturé par des poissons carnassiers, le cycle continue. Si le copépode est ingéré par un poisson non carnassier, le cycle est bloqué et ne pourra reprendre que si la larve se retrouve finalement dans un poisson carnassier. Dans l’estomac du poisson carnassier, les larves entrent dans la paroi et vont se loger dans la musculature (surtout dans les flancs). Du stade procercoïde, on passe ou stade plérocercoïde qui est infestant pour l’Homme. Si cette dernière larve est mangée par un hôte définitif, elle donnera le stade adulte assez rapidement.
Dans certains cas, le poisson carnassier peut être mangé par un autre poisson : les larves pleurocercoïdes ne restent alors pas dans l’estomac mais partent dans les viscères du dernier poisson. Là, elles pourront survivre pendant plusieurs années, sans grandir ni se différencier. Le cycle continue si ce poisson est mangé par un poisson carnassier…
La Ligule intestinale : Ligula intestinalis. La ligule adulte se trouve dans l’intestin d’oiseaux aquatiques piscivores (comme les cormorans ou les canards). Le premier hôte intermédiaire est un crustacé copépode. Le deuxième hôte intermédiaire est un poisson cyprinidé. C’est dans la cavité abdominale de ces poissons que les larves se rencontrent. Elles peuvent atteindre plus de 25 centimètres de long et il peut y avoir plusieurs larves dans un même poisson. Il va se produire une déformation importante de la partie ventrale : le corps s’alourdie et la capacité de nage s’en trouve diminuée. Ces poissons deviennent plus faciles à attraper par les oiseaux.
Ce parasitisme entraîne une stérilisation des poissons car il n’y a plus d’espace libre pour le développement des gonades.

Ce tænia vit dans l’intestin grêle. Sa forme larvaire est localisée dans le cerveau de mouton (c’est la forme cénure) dont la taille est celle d’un œuf de poule (6 à 8 centimètres). La présence de cette larve provoque des troubles de la mobilité : c’est la maladie du tournis.
C’est sur la forme larvaire que vont bourgeonner de nombreux scolex, au niveau de la membrane proligère. La formation de nombreux scolex s’apparente à une multiplication asexuée.
Tænia échinocoque : Echinococcus granulus.

Le vers traverse la paroi intestinale et rejoint le foie de l’herbivore et là, se différencie en une boule de 20 centimètres de diamètre : le stade hydatide. Ce stade se développe lentement : 16 mois chez les ovins et les bovins ; 18 mois à 30 ans chez l’Homme. Dans l’hydatide, il y a un bourgeonnement de scolex à partir de la membrane interne (proligère) : on peut arriver à 400 000 scolex par centimètre cube. Chez l’Homme, il y a besoin d’une intervention chirurgicale. Les plus grands foyers de développement sont les lieux d’élevage intensif de mouton.
Echinococcus multilocularis (Echinococcus alveolaris).
Ce vers est un parasite du renard (hôte définitif). L’hôte intermédiaire est le campagnol. L’Homme peut être hôte intermédiaire s’il consomme des baies sauvages souillées. Il existe une forme hydatide (dans le foie) qui provoque des nécroses hépatiques très graves.
Chez l’Homme, cette maladie est l’échinococcose alvéolaire ; 200 cas étaient recensés en France, vers 1980.
Diphylidium cannium (cyclophyllidés).
C’est un vers dont la taille varie entre 50 et 80 centimètres. L’adulte est dans l’intestin de chien. L’œuf se développe quand il est avalé par des larves de puces. La puce subit des mues ou des métamorphoses et le chien se contamine en ingérant des puces adultes.
II Les Pseudocœlomates.
Les pseudocœlomates ont leurs organes situés dans une cavité corporelle qui a des avantages réels. Celle-ci est appelée cavité viscérale ou primaire. Toutefois, le mésoderme ne participe que partiellement à la délimitation de cette cavité (du coté externe uniquement).La cavité renferme quelques cellules amiboïdes. Le tube digestif et les gonades sont suspendus dans cette cavité.
On définit sept embranchements dont trois qui seront étudiés : les Nématodes, les Nématomorphes et les Rotifères. Anciennement, les nématodes et les nématomorphes composaient le groupe unique des némathelminthes.
Les pseudocœlomates ont tous un plan d’organisation et des caractères semblables :
- Ils sont tous plus ou moins vermiformes.
- Leur corps est recouvert d’une cuticule.
- La paroi du corps ne possède pas de couches musculaires circulaires et transversales.
- La paroi du tube digestif est généralement simple (cellules épithéliales sur une couche) avec parfois une couche musculaire autour de la paroi.
- Il n’y a ni système respiratoire ni système circulatoire.
- Le système excréteur est le plus souvent protonéphridien.
- Les pseudocœlomates présentent un cloaque où débouchent les systèmes digestif, urinaire et génital.
La plupart des pseudocœlomates présente un nombre défini de cellules dans chaque organe. C’est l’eutélie. La croissance se fait grâce à l’augmentation du volume cellulaire et non du nombre de cellules. On pourra fréquemment observer des cellules géantes.
Toutefois, cette tendance limite les possibilités de régénération. En fait, cette tendance a été étudiée chez Cœnorhobditis elegans (génétiquement très connu, comme la drosophile) et il a été montré un même complexe de gènes homéotiques que chez la drosophile, la souris et l’Homme (HOM/HOX).
A Les Nématodes (semblables à un fil).
Chez les nématodes, on observe l’absence totale de cellule flagellée ou ciliée (même chez les spermatozoïdes). Les spermatozoïdes sont ronds ou coniques et se déplacent grâce à des mouvements amiboïdes.Il existe plusieurs dizaines de milliers d’espèces (80 000) mais elles sont difficiles à identifier. Les nématodes sont souvent parasites de végétaux ou d’animaux (dont l’Homme) et provoquent des nématodoses (dangereuses).
D’un point de vue écologique, on les trouve sous toutes les latitudes (des glaciers jusqu’aux sources thermales) et ils présentent une grande uniformité structurale.
1 Morphologie.
Les nématodes sont des vers blancs à section circulaire, effilés aux deux extrémités. Les mâles ont une taille comprise entre 12 et 15 centimètres alors que les femelles sont bien plus grandes. La bouche s’ouvre à l’extrémité antérieure et est entourée de trois lèvres (une dorsale et deux ventrales) portant des soies céphaliques à rôle sensoriel.La femelle a une extrémité postérieure qui porte l’anus. Le mâle a une extrémité postérieure enroulée ventralement, en crosse, et qui porte, en position sub-terminale, un orifice sub-cloacal où l’on trouve des spicules sexuels copulateurs.
La femelle présente une constriction annulaire dans le tiers antérieur du corps et c’est à ce niveau que s’ouvre l’orifice génital ventral.
Lors de l’accouplement, le mâle enroule sa crosse autour de la femelle, dans le tiers antérieur.
2 Tégument.

Sous la cuticule se trouve l’épiderme avec deux épaississements longitudinaux (un dorsal et un ventral) où sont localisés les cordons nerveux (un dorsal et un ventral). Latéralement (à gauche et à droite) on trouve deux gouttières au niveau desquelles on a les canaux excréteurs.

Ces cellules sont sur une assise et forment quatre champs musculaires (deux dorsaux et deux ventraux). Ces champs sont délimités par quatre cordons (un dorsal, un ventral, un à droite et un à gauche). Ces cellules différencient des myofibrilles du côté de l’épiderme : c’est un dispositif original car d’habitude, ce sont les nerfs qui envoient des fibrilles vers les organes. La contraction de ces cellules donnera uniquement des enroulements.
3 Les organes des sens.
Il existe des organes classiques mais aussi :- Des Amphides : invaginations cuticulaires dans la partie antérieure à rôle chimiorécepteur.
- Les Phasmides : ce sont des organes sensoriels paires, situés latéralement en position post-annulaire.
4 Le système nerveux.
Le système nerveux des nématodes est rudimentaire, acculé ou inclus dans l’épiderme. Il est composé de centres nerveux localisés au niveau du collier péri-œsophagien. Ce collier émet des nerfs en direction des lèvres buccales et va émettre des troncs nerveux moteurs ou sensitifs. On distingue six nerfs dans la région antérieure et huit troncs nerveux (6 moteurs et 2 sensitifs) qui partent vers la partie postérieure.5 L’appareil excréteur.
L’appareil excréteur ne porte pas de protonéphridie. On a deux types distincts :Chez les nématodes primitifs, on a une ou deux cellules géantes (les cellules Renette) situées ventralement à la jonction œsophage/intestin. Chaque cellule possède un pore excréteur en position antérieure.
Chez les formes plus évoluées, les cellules géantes s’atrophient et il y a formation de deux canaux excréteurs qui courent le long du corps ; un seul pore excréteur est présent en position antérieure.
6 L’appareil digestif.
L’appareil digestif est pratiquement rectiligne et comporte cinq éléments principaux :- La bouche (en position terminale avant) est entourée par trois lèvres hémisphériques (une dorsale et deux ventrales).
- La cavité (ou capsule) buccale est tapissée par une mince couche cuticulaire ; l’organisation est fonction du comportement alimentaire des différentes formes. Cette capsule buccale peut être armée de dents, denticules ou râpes. Chez les nématodes marins les plus primitifs, quatre formes existent :
- Forme simple sans armature : elle permet l’absorption de nourriture liquide.
- Forme conique sans armature : on la trouve chez les nématodes microphages.
- Forme à armature réduite (denticules ou râpes) : elle est présente chez les mangeurs d’algues ou chez certaines formes mégalophages (qui mangent de tout).
- Forme à armature développée : existe chez les nématodes prédateurs.
- Le pharynx est musculeux à parois épaisses, tapissé d’une couche cuticulaire. C’est à ce niveau que l’on trouve des glandes à fonction anti-coagulantes (pour les formes parasites de vertébrés).
- L’intestin est droit et mince (une seule couche cellulaire endodermique).
- Le rectum est d’origine ectodermique car il est tapissé d’une couche cuticulaire. Les femelles ont un rectum terminé par un anus qui s’ouvre en position sub-terminale. Chez le mâle, le rectum débouche dans un cloaque où l’on trouve les spicules copulateurs. C’est ici que s’ouvrent les canaux déférents.

D’abord, la partie antérieure se dilate : il y a aspiration de liquide; la valve terminale est fermée.
Ensuite, le pharynx se contracte : la partie antérieure va avoir un diamètre plus petit ; la valve postérieure s’ouvre et le liquide passe par cette valve.
7 L’appareil génital.

a L’appareil mâle :
Cet appareil est constitué d’un testicule tubulaire, filiforme, très long (1 mètre ou plus). Il est entouré sur lui-même et autour de l’intestin. Ce testicule se renfle dans la portion terminale et forme la vésicule séminale qui stocke les spermatozoïdes. Cette vésicule débouche dans le cloaque.b L’appareil femelle :
L’appareil femelle est composé de deux ovaires tubulaires filiformes (un à deux mètres). Ces ovaires se transforment ensuite en oviductes puis en utérus bien développés. Ils vont se rejoindre pour donner le vagin qui est beaucoup plus grêle. Il aboutit à l’orifice génital femelle (dans le tiers de la partie antérieure, ventral).Les femelles fécondées pondent de 70 000 à 240 000 œufs par jour pendant 12 à 18 mois.
La fécondation est toujours croisée et interne grâce aux stylets copulateurs du mâle.
Les œufs sont pondus à différents stades selon l’espèce. La ponte peut donner :
- Des œufs non segmentés.
- Des œufs à segmentation débutée.
- Des œufs à embryon complètement formé.
La cuticule externe oblige l’accroissement à se faire par mues successives (en général, il y a quatre mues dans la vie d’un animal).
8 Les grands groupes de nématodes.
On distingue deux principaux groupes :- Les Aphasmidiens : ils n’ont pas de phasmide et sont parasites.
- Les Phasmidiens : ils possèdent deux phasmides et sont libres, vivant dans la terre ou dans la matière putréfiée ; sinon, ils peuvent être parasites de végétaux ou d’animaux.
9 Ecologie et éthologie des formes libres.
Les formes libres représentent la moitié des nématodes. On les rencontre dans la plupart des habitats (eau douce, eau de mer, sol humide, mousse, matière organique en décomposition).Souvent, les nématodes pullulent :
- Dans les fruits pourris des vergers : on trouve plusieurs milliers d’individus d’espèces différentes.
- Dans les milieux hostiles au développement de la vie. Exemple : l’anguille du vinaigre (Anguillula) se développe dans le vinaigre, avec 6 à 7% d’acide acétique pur. Elle se nourrit du cadavre des bactéries acétiques, responsables de la fermentation. Elle supporte des pH allant de 2,5 à 11,5.
10 Ecologie et éthologie des formes parasites.
Généralement, en cas de parasitisme, il y a des modifications structurales importantes et présence d’une reproduction asexuée possible à l’état larvaire.Ces modifications ne se voient pas chez tous les nématodes. Le développement des formes libres ou parasites est identique. Les parasites vont s’attaquer aux végétaux ou aux animaux.
a Les parasites de végétaux.

b Les parasites d’animaux.
Exemple 1 : Enterobius vermicularis.
Ce parasite est responsable de l’oxyurose.Les adultes vivent et s’accouplent dans le cæcum digestif de l’Homme, pouvant provoquer l’appendicite. Après accouplement, les femelles vont dans le rectum, s’y fixent et pondent. Il y a alors démangeaisons ou prurit anal. Les œufs (de 50 à 60µm) restent collés dans l’anus. Ils donnent les embryons qui sont directement infestant.
L’enfant se contamine en portant les mains (souillées) à la bouche (c’est une auto-infestation ou parasitose des mains sales), ou en inspirant des poussières contenant des œufs.
Les œufs, une fois dans le tube digestif vont subir l’action des sucs digestifs qui vont casser la coque et ainsi, libérer les larves qui passent au stade adulte en une trentaine de jours. Les adultes se nourrissent des bactéries du tube digestif et ne sont pathogènes que lorsque leur population augmente trop fortement et provoque des occlusions du tube digestif.
Exemple 2 : Trichurus trichuria (monoxène).
Ces vers sont des parasites des cæcums digestifs. Ils ont une partie antérieure effilée, implantée dans la muqueuse digestive des mammifères (la partie postérieure est libre). Ce sont des hématophages. Une centaine d’individus consomment 50 millilitres de sang par jour.Les œufs pondus ne donnent pas directement des larves (en restant à l’intérieur). Pour la suite de leur développement, un séjour de trois semaines à l’extérieur est obligatoire.
La durée de vie de l’adulte est de cinq ans. Les femelles donnent 10 000 œufs par jour et l’infestation se fait par l’intermédiaire de boissons et de légumes mal lavés.
Les œufs donnent des adultes en un mois.
Exemple 3 : Ascaris.
Les larves d’ascaris font d’importants déplacements dans l’organisme parasité : migration dans l’hôte (à cycle avec migration).Les lombricoïdes provoquent l’ascaridiose humaine.
Les adultes s’accouplent et pondent dès l’intestin grêle. La femelle donne 240 000 œufs par jour. Ces derniers seront évacués avec les excréments et donnent des larves infestantes au bout de 30 à 40 jours si la température externe est voisine de 30 degrés, avec des sols humides et ombragés.
L’infestation est réalisée par ingestion d’aliments souillés. Après la digestion de la coque, la larve va migrer dans l’organisme. Les larves traversent d’abord l’épithélium digestif avant d’atteindre le foie par voie sanguine. Elles y restent pendant trois à quatre jours puis, gagnent le cœur droit par la circulation veineuse puis rejoignent les poumons par les artères pulmonaires et y restent environ une semaine. Elles vont y subir deux mues. Elles remontent ensuite les voies respiratoires (bronchioles, bronches, trachée) et arrivent au niveau du carrefour bucco-pharyngé et prennent alors la voie digestive et rejoignent le grêle où elles deviendront adultes. La migration dure environ 15 jours. Après le grêle, les larves passent au gros intestin, puis au rectum où elles sont matures sexuellement.
L’homme est le seul réservoir de ce parasite et cette maladie touche environ ¼ de la population mondiale, surtout dans les pays pauvres.
Les symptômes observés sont : des occlusions intestinales, des invaginations intestinales (et des étranglements de ce tube digestif à hernies), des péritonites aiguës et des dégâts dans le foie (pancréatites). De plus, ces ascaris rejettent des substances toxiques pour le système nerveux de l’Homme. D’autres vertébrés or l’Homme peuvent être parasités par pratiquement la même espèce (cheval, porc, chien, chat).
Exemple 4 : Trichinella spiralis (hétéroxène).
On trouve ce parasite dans l’intestin grêle de nombreux animaux (chien, chat, rat, porc, sanglier, phacochère, Homme). Les femelles pénètrent dans la muqueuse intestinale et y pondent deux larves par heure pendant les vingt-huit jours de leur vie.Les femelles sont vivipares et donnent des larves qui gagnent le cœur droit par le liquide lymphatique puis, les poumons, passent dans le cœur gauche et seront distribuées dans tout l’organisme par la circulation aortique. Ces larves vont se fixer dans les muscles striés et s’enkyster pendant 12 à 18 mois. Les kystes se calcifient et sont infestant. Les Hommes se contaminent en consommant de la viande mal cuite.
Chez les rats, l’infestation est propagée par le cannibalisme caractérisant cette espèce : l’hôte intermédiaire et l’hôte définitif peuvent être de la même espèce. Dans ce cas, on parle alors de cycle auto-hétéroxène.
Exemple 5 : Wuchereria bankrofti (Filaire de Bancroft).
Les adultes de ce parasite vivent dans les vaisseaux lymphatiques. Le mâle fait environ 4 centimètres et la femelle mesure aux alentours de 8 centimètres. Les larves circulent dans le liquide lymphatique et migrent périodiquement dans les vaisseaux sanguins périphériques selon un rythme nycthéméral (jour/nuit) correspondant à la sortie de l’insecte vecteur (Anophèle : moustique). Grâce aux piqûres, les moustiques pompent le sang de vertébrés alors que les larves se concentrent sur les pièces buccales du moustique, Elles seront transmises à la victime suivante par une nouvelle piqûre. Les larves deviennent adultes en trois mois. Les parasites sont surtout dans les zones tropicales (Afrique, Amérique du sud, Asie).Ce parasite provoque l’éléphantiasis : les membres inférieurs deviennent énormes à cause de la présence du parasite dans le système lymphatique de ces membres. Le scrotum peut aussi être atteint.
B Les nématomorphes.
Les nématomorphes sont des vers longs (environ 1,5 mètre) qui ressemblent aux nématodes. Leur intestin est réduit, les larves sont parasites alors que les adultes sont aquatiques libres.La vie adulte est brève et ces adultes ne se nourrissent pas : l’anus et la bouche sont non-fonctionnels. La larve va donc se nourrir par osmose, au travers du tégument.
Le système nerveux est intra-épidermique ; canaux excréteurs et protonéphridies sont absents ; on note la présence de cellules myo-épithéliales sur un seul champ ventral. Les sexes sont séparés ; il y a un dimorphisme sexuel ; la fécondation est interne.
On trouve environ 250 espèces de nématomorphes, regroupées en deux ordres : les Gordiens (larves parasites d’insectes en eau douce) et les Nectonématoïdes (parasites de crustacés, marins).
1 Les Gordiens.
Chez les gordiens, le pseudo-cœlome est pratiquement rempli par du mésenchyme.Exemple : Paragordius tolosanus.
Ce parasitepossède deux hôtes intermédiaires.Les œufs donnent des larves qui vont au fond de l’eau. Elles seront ingérées par une larve d’insecte névroptère (Sialis) qui doit être mangée par un carabe (coléoptère). Une fois dans ce carabe, la larve se métamorphose en un vers adulte, libre. Ce dernier pourra réaliser la fécondation.
2 Les Nectonématoïdes.
Les nectonématoïdes sont parasites de crustacés. Les adultes sont libres et pélagiques ; mesurent environ 20 centimètres de long. On note la présence d’une double rangée de soies sur le corps. Ces soies favorisent la nage par ondulation. Le pseudo-cœlome n’est pas comblé par du mésenchyme.Exemple : Nectonema.
C Les Rotifères.
Les rotifères ont une taille généralement inférieure à celle des ciliés (ils dépassent rarement deux millimètres). Leur corps est non-segmenté et la partie antérieure du corps est pourvue d’un appareil rotateur cilié (caractéristique de cet embranchement).1 L’appareil rotateur.
Cet appareil est constitué par un ou deux anneaux de cils entourant une zone centrale déprimée en entonnoir (l’aire buccale). Au fond de cette aire s’ouvre la bouche. Cet appareil sert à la locomotion et à la nutrition. Il est très développé, ou réduit, ou absent, pour des formes de rotifères semi-terrestres.La couronne extérieure postérieure est appelée paratroque (ou cingulum). La couronne interne antérieure est appelée prototroque ou trochus.
Sur la face dorsale, on peut trouver des tentacules garnis de soies sensorielles tactiles (le nombre de tentacules est fonction de l’espèce).

2 Le pied.
La région postérieure est appelée « le pied ». Celle-ci est terminée par deux orteils plus ou moins longs entre lesquels débouchent les glandes pédieuses qui sécrètent un liquide visqueux.Chez certaines espèces, les orteils forment des ventouses ou des disques adhésifs qui servent aux déplacements ou à la fixation de l’animal un substrat dur.
Chez les espèces pélagiques, le pied peut être réduit ou absent. Il est très développé chez les formes primitives qui sont rampantes.
3 Paroi et musculature.

L’épiderme du pied (glandes pédieuses) sécrète une substance pour la synthèse de tubes chez les rotifères fixés. Cet épiderme donne chez les formes pélagiques, une substance gélatineuse.
La musculature est essentiellement constituée de faisceaux circulaires, longitudinaux et discontinus. Il existe des muscles rétracteurs du pied et de l’appareil rotateur.
Il existe un organe rétro-cérébral dans la région antérieure (au niveau du système nerveux central) qui a la forme d’un sac ouvert à l’extérieur. La fonction de cet organe est inconnue.
4 L’appareil digestif.
La bouche s’ouvre au fond de l’aire buccale (antérieure). Celle-ci communique avec un pharynx de deux manières :- Directement chez les espèces prédatrices.
- Par l’intermédiaire d’un tube buccale à paroi interne ciliée.
On peut distinguer sept pièces cuticulaires principales : le fulcrum, les ramus (x2), les uncus (x2), les manubrium (x2).
Les ramus et le fulcrum constituent la mâchoire inférieure alors que les uncus et les manubriums constituent la mâchoire supérieure. Chez les espèces prédatrices, cet appareil peut faire saillie par l’orifice buccal pour permettre la capture et la dilacération des proies. Les pièces de cet appareil ont une morphologie différente, comme la formation des pinces ou des pièces perforantes.
Au niveau du pharynx, on trouve des glandes salivaires (entre deux et sept en fonction des espèces). L’œsophage est court et cilié. L’estomac est glandulaire ; on y trouve deux glandes gastriques. L’intestin est court, suivi d’un rectum qui débouche dans un cloaque. L’orifice cloacal est dorsal, au niveau de la descente du pied.
5 L’appareil excréteur.
L’appareil excréteur est formé par deux protonéphridies. Les canaux en partant débouchent dans une vessie contractile (ou urocyste). Elle débouche ventralement dans le cloaque. Chez certaines espèces, la vessie est absente : ce sont alors les canaux qui débouchent dans le cloaque. Cet appareil contrôle la pression interne et joue donc un rôle osmo-régulateur.6 Système nerveux et organes des sens.
Le système nerveux est composé par un volumineux ganglion cérébral. De ce ganglion partent deux nerfs ventraux et deux terminaisons nerveuses secondaires vont innerver l’appareil rotateur, les yeux, la musculature et partent vers le corps.Les organes sensoriels se situent au niveau de la double couronne ciliaire de l’appareil rotateur. On distingue trois types d’organes : les chémorécepteurs, les tangorécepteurs (rôle tactile), les photorécepteurs.
Les organes sensoriels sont : des soies, des papilles, des fossettes ciliées. On trouve aussi deux yeux qui vont fusionner en un œil unique rouge vif.
7 L’appareil reproducteur.
Les sexes sont séparés (gonochorisme) et le dimorphisme sexuel est très marqué (les mâles sont plus petits que les femelles). Pour de nombreuses espèces, la forme mâle n’a jamais été découverte.L’appareil mâle:
Cet appareil est constitué d’un testicule (donc impair) poursuivit par un canal déférent qui s’ouvre au niveau du pied. Il peut y avoir formation d’une papille à l’ouverture, représentant un pénis.L’appareil femelle:
Cet appareil est formé d’un ovaire en position ventrale, suivie d’un oviducte qui débouche dans le cloaque. L’ovaire est composé de deux parties : la partie antérieure comprend les ovules alors que la partie postérieure à un rôle vitellogène. On observe donc une séparation des fonctions gamétogènes et vitellogènes.8 Systématique.
Les Rotifères sont divisés en trois ordres :- L’ordre des Seisonidés. Dans ce groupe, on trouve quelques espèces marines dont certaines qui sont épizoïques (vivant fixées sur d’autres organismes). Leur reproduction est bisexuée, sans parthénogenèse.
- L’ordre des Bdelloïdés. Les bdelloïdés colonisent les mousses, les lichens d’eau douce. Ils sont nageurs ou rampants. On ne connaît pas les formes mâles et il y a parthénogenèse thélytoque (femelle à femelle), indéfinie et obligatoire.
- L’ordre des Monogonontes. Les monogonontes vivent dans les eaux douces, saumâtres, dans les mousses, les lichens. Ils peuvent être nageurs ou fixés.
9 Mode de vie.
Actuellement, on connaît environ 2000 espèces. On les trouve dans les différents milieux de vie mais particulièrement en eau douce et dans les mousses et lichens. On note quand même la présence de quelques formes parasites de végétaux et d’animaux.Les formes aquatiques sont surtout benthiques et se déplacent à la recherche de nourriture. Les autres formes sont fixées (généralement sur des algues). Les formes planctoniques ont une morphologie externe modifiée par les conditions du milieu.
Pour une même espèce, on va avoir une morphologie différente selon le milieu de vie, ce qui entraîne des difficultés de détermination.
Les rotifères mangent des animaux, des végétaux (souvent de petite taille) qui seront attirés par la (les) couronne(s) ciliaire(s). Le mastax, chez les formes
prédatrices est la première manifestation de la fragmentation des aliments dans le tube digestif.
10 Reproduction.
Les œufs ont une segmentation totale et égale.Chez les Seisonidés, la reproduction est simple, normale. Il n’y a jamais de parthénogenèse. Chez les Bdelloïdés, toutes les femelles donnent des œufs qui se développent seulement par parthénogenèse. En revanche, les Monogonontes ont une reproduction sexuée compliquée (avec trois sortes d’œufs). Il y a alternance de générations parthénogénétiques et d’une génération sexuée.
Les femelles parthénogénétiques sont appelées « femelles amictiques ». Elles sont diploïdes et ne peuvent s’accoupler. Elles donnent des embryons à développement immédiat ne subissant pas la réduction chromatique. Ces embryons donnent des femelles diploïdes. Sous l’influence de certaines conditions du milieu (photopériode, température, pH, densité de population) apparaissent des femelles fécondables (« femelles mictiques »). Ces dernières vont donner des ovules qui émettent deux globules polaires. Si les ovules sont non fécondés, on aura des mâles haploïdes. Ces mâles pourront s’accoupler avec les femelles mictiques. La fécondation aboutit à des œufs à coquille épaisse à « œufs de durée ». Ces œufs peuvent supporter l’hiver. Quand ils se trouvent dans de bonnes conditions, ils se développent et produisent des femelles amictiques.
Les rotifères peuvent être dits monocycliques quand ils présentent plusieurs générations parthénogénétiques pendant la même saison et une génération sexuée à l’automne. Les rotifères polycycliques ont au moins deux périodes de reproduction sexuée (automne et printemps).
Pour tous, à la belle saison, il y a succession de femelles amictiques (diploïdes).
Il existe des affinités nématodiennes : symétrie bilatérale, pseudo-cœlome, cuticule, absence d’appareil respiratoire et circulaire.
Il existe aussi des différences fondamentales avec les mêmes nématodes : les nématodes ont une croissance discontinue par mues, ils n’ont pas de formations ciliaires ni d’appareil excréteur de type protonéphrien.
Nématodes et rotifères n’ont donc pas d’ancêtre commun.
Par contre, on trouve des ressemblances remarquables avec les plathelminthes. Le système excréteur est proche, il y a séparation des fonctions gamétogènes et vitellogènes. On trouve les mêmes modalités de développement des rotifères et des turbellariés. Il y a la présence d’une ciliature assurant la locomotion. On peut aussi comparer les pièces du mastax et les pièces du pharynx de certains turbellariés.
Une parenté entre turbellariés et rotifères est donc probable.