
I Métabolisme.

A Le métabolisme glucidique.
1 Le glucose.
a La formation du pyruvate.
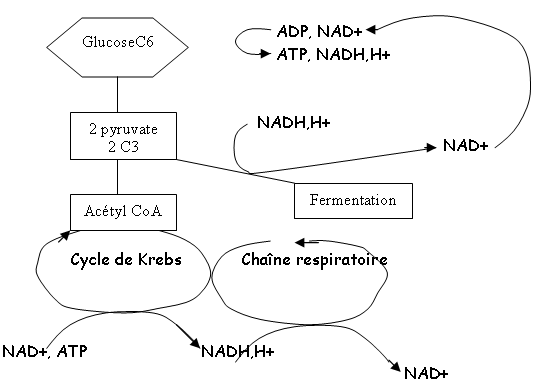
• La glycolyse1 C6 → 2 C3
Bilan : 2 ATP + 2 NADH,H+ (le NADH,H+ est présent pour récupérer les électrons au cours des réactions rédox.
• La voie du KDPG
C’est une voie alterne à la glycolyse.
1 C6 → 2 C3
Bilan net : 2 NADH,H+ + 1 ATP.
• La voie des pentoses-phosphates.
Cette voie est complémentaire à la glycolyse. Elle a un rôle de dégradation des sucres en C5 pour former des C4.
Bilan : 3 C6 → 3 C5 + 3 CO2 + 9 NADH,H+.
b Le devenir du pyruvate.
On a cinq possibilités :- fermentation : le pyruvate devient un substrat (il peut être réduit en acide lactique). La respiration se combine avec le coenzyme A qui passe dans le cycle de Krebs.
- La respiration aérobie : il y a production d’ATP et accumulation de NADH,H+. Ces deux composés vont aller alimenter les chaînes respiratoires (dans les mitochondries ou dans les membranes bactériennes) et régénérer du NAD+. NADH,H+ → NAD+ ; O2 → H2O ; 1 C6 → 6 CO2. On a un bilan total de 38 ATP pour les procaryotes et de 36 ATP pour les eucaryotes (2 ATP sont utilisés pour traverser la paroi mitochondriale). Et le NAD+ est régénéré.
- La respiration anaérobie : l’accepteur final peut être différent type de molécules. On peut trouver des nitrates (NO3- → N2), des sulfates (SO42- → H2S), CO2 → CH4. Ce sont des chaînes incomplètes : le bilan donne toujours 6 CO2 mais moins de 38 ATP et moins de NAD.
- Les fermentations : leur but est de régénérer le NAD+. Elles ont le même bilan que la glycolyse. 1 C6 → 2 C3 Acide lactique → → + 2 ATP→ → + 2 NADH,H+ 2 NAD+
B Les Polysaccharides.
La filière générale permet de dégrader les oses en glucose. Si on obtient un autre composé, ce sera du fructose qui rentrera plus loin dans le cycle.La cellulose : les enzymes cellulases sont fabriquées par des micro-organismes de type Actinomycètes, Bacillus, Vibrio, Cytophage.
C Le catabolisme protéique.
On trouve les micro-organismes responsables de ce catabolisme dans les putréfactions et les gangrènes. Il y a hydrolyse des protéines pour former des amines volatiles qui donneront les mauvaises odeurs. Il y a ensuite une phase d’ammonisation : les acides aminés donnent des acides organiques quelconques et de l’ammoniac.D Le catabolisme des lipides.
Ils sont dégradés en acétylCoA qui va rejoindre le cycle de Krebs.• Nocardia permet la dégradation des pneus.
• Certaines bactéries stimulées vont être capables de dégrader les nappes de pétrole.
II Génétique.
C’est la recombinaison et les transferts génétiques. Ces deux phénomènes ont un rôle de reproduction sexuée chez les micro-organismes qui n’en ont pas naturellement. Les recombinaisons ne touchent que les secteurs homologues.A Les mutations.
Une mutation est un changement spontané. Il y a une mutation pour 10^6 gènes répliqués. On obtient alors une bactérie mutante toutes les 10^7 bactéries formées. Parmi les facteurs mutagènes, on trouve les UV, les radiations…B Les recombinaisons (parasexualité).
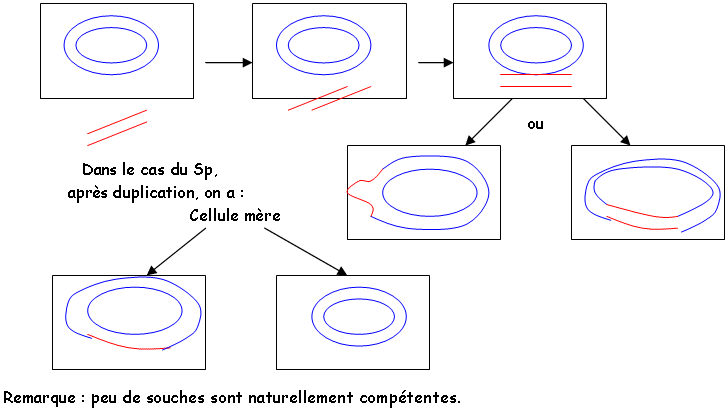
Les recombinaisons ont lieu grâce à trois mécanismes (peut-être quatre) et entraînent la formation d’un mérozygote (diploïde partiel)1 La transformation (l’expérience de Griffith).
On travaille à partir de Streptococcus pneumoniae sous deux souches : virulente (SpE) et non-virulente (SpN).Si on chauffe SpE et qu’on l’injecte dans une souris, la souris ne meurt pas. Si l’on réalise un broyat de SpE avec des SpN et qu’on injecte celui-ci dans la souris, elle meurt.
En 1944, Avery, McLeod, McCarthy découvrent la solution à ce problème : l’ADN.
Une bactérie intacte associée à un fragment d’une autre entraîne une transformation. Ce phénomène demande une «compétence » de la cellule réceptrice. Cette compétence s’acquière en fin de phase de multiplication de la cellule (faculté de perméabilité).
On a trois étapes : compétence, synthèse d’ADNliase, synthèse d’ADNhydrolase.
2 La transduction.
Ce phénomène est en relation avec des virus.a La transduction généralisée.
Un phage arrive, injecte son ADN sans hydrolyser le génome bactérien : on a alors un virus avec ADN bactérien.b La transduction spécialisée.
Ce phénomène est réalisé par un phage tempéré. Il y a excision de l’ADN du phage du génome bactérien. Ce sont toujours les mêmes gènes qui sont concernés.3 La conjugaison.
C’est un échange d’ADN par contact entre bactérie réceptrice et donatrice. L’ADN concerné est souvent le plasmide F. On parle pendant la conjugaison, de bactérie + (mâle) et de bactérie – (femelle). L’adhésion est permise grâce aux F-pili ou pili sexuels.Le canal de conjugaison est encore mal connu. L’ADN échangé est de l’ADN monobrin.