Introduction.

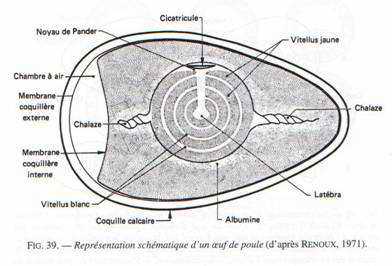
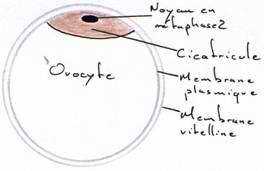
Au niveau de l’ovaire : l’ovocyte s’entoure de couches concentriques de vitellus, lequel est limité par une membrane vitelline. Le jaune représente l’ovocyte proprement dit. Le noyau est rejeté à la périphérie de ce vitellus et se situe dans une région périplasmique qui a l’aspect d’un disque : la cicatricule.
La vésicule germinative représente le noyau en prophase 1.
Ce vitellus permet le développement embryonnaire pendant 21 jours. Il est riche en lipides et fortement coloré par les caroténoïdes (coloration rouge). Durant son transit dans l’oviducte et après sa fécondation, le vitellus est entouré par l’albumen (de nature protéique : le blanc). Ce blanc représente pour l’embryon un supplément de réserves. Il est entouré par une membrane coquillaire double qui s’écarte au gros bout de l’œuf pour constituer la poche à air. Dans la partie inférieure de l’oviducte, l’œuf est revêtu d’une coquille calcaire.
Seul l’ovaire gauche est fonctionnel.
La libération de l’ovule de l’ovaire ou ponte ovulaire provoque la reprise de la première division méiotique et donne un ovocyte 2 bloqué en métaphase 2.
La fécondation est interne et se fait dans la partie antérieure de l’oviducte, avant la sécrétion de l’albumen dans les 15 minutes qui suivent l’ovulation. Il y a polyspermie ; la pénétration du spermatozoïde provoque l’activation de l’œuf et la reprise de la 2ème division méiotique. Le développement embryonnaire commence durant le transit dans l’oviducte. La segmentation dure 24 heures puis, s’achève au moment de la ponte. Ce développement ne reprendra que si l’œuf est incubé à 38°C.

I La segmentation.
La segmentation va être partielle compte tenu de la taille, qui se limite à la cicatricule (disque au niveau du pôle animal). Cette segmentation commence 5 heures après la fécondation. Le premier plan de segmentation clive la cicatricule en deux et donne 2 blastomères.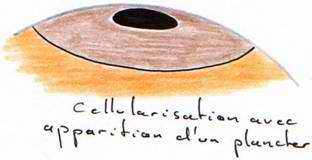
Le second plan de segmentation est aussi superficiel, perpendiculaire au premier et donne 4 blastomères. Ces deux plans de segmentation peuvent être considérés comme des méridiens. Le troisième plan se fait selon la verticale, perpendiculaire au second et donne huit blastomères. Le quatrième site de segmentation se fait selon la verticale, englobant huit blastomères au centre et huit à la périphérie : on atteint alors un stade de 16 blastomères.
Des plans horizontaux vont fermer ces blastomères. L’ensemble de la cicatricule ainsi segmentée est le blastoderme (64 à 128 blastomères).

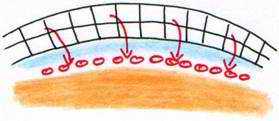
Une cavité se creuse, le blastocoele primaire qui donnera la blastula primaire avec deux régions :
- l’aire pellucide donnera l’embryon proprement dit
- deux aires latérales : les aires opaques donnant la partie extra-embryonnaire.
Ces deux feuillets délimitent une cavité qui est le blastocoele secondaire, équivalent du blastocoele de la blastula des amphibiens. Entre l’entophyle et le vitellus, il y a une cavité appelée l’archantéron primaire.
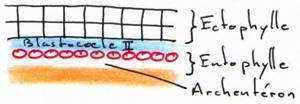
A ce stade, la symétrie bilatérale de l’embryon est acquise. Le développement s’arrête s’il n’y a pas incubation après la ponte de l’œuf.
II Développement durant le début de la gastrulation.
Juste avant le début de la gastrulation, une lame basale va se constituer sous l’ectophyle puis, des cellules issues de l’ectophyle vont migrer en profondeur et constituer l’hypoblaste et l’entophyle. L‘hypoblaste et l’entophyle donneront ultérieurement l’endoderme extra-embryonnaire qui recouvrera la totalité du vitellus.III La gastrulation.
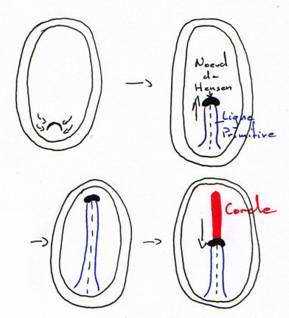
Pendant la migration, on voit apparaître un soulèvement.La gastrulation débute par un épaississement de l’aire pellucide dans la région postérieure. C’est dû à la convergence des territoires superficiels vers la région médiane et postérieure. Cet épaississement s’allonge vers la 6ème heure d’incubation. Le blastoderme prend une forme ovale.
Entre la 10ème et la 16ème heure d’incubation, cet épaississement prend le nom de ligne primitive qui se creuse d’un sillon (fosselle primaire) qui correspond au blastopore des amphibiens. Elle est limitée vers l’avant par une région plus compacte : le nœud de Hensen qui est l’équivalent de la lèvre dorsale du blastopore (responsable de la mise en place de corde).
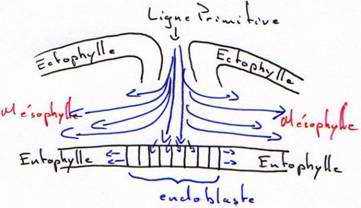
On a pu reconstituer une carte des territoires présomptifs (en 1937, grâce à Pasteels) par un marquage des cellules du blastomère par les particules d’encre de Chine qui permet d’analyser les mouvements morphogénétiques et la destinée des différents feuillets.
Si on se réfère à cette carte, on voit, qu’au fur et à mesure de l’allongement de la ligne primitive, c’est le matériel superficiel du mésoblaste des lames latérales et extra-embryonnaires qui converge le premier puis immigre en profondeur et diverge pour donner une lame mésodermique pleine, qui va s’insinuer entre l’ectophyle et l’hypoblaste.
Entre 10 heures et 16 heures, le mésoblaste précordal passe en profondeur au niveau du nœud de Hensen. Ce mésoblaste cordal converge vers l’axe médian puis s’insère entre l’ectophyle et l’entophyle puis s’allonge vers l’avant pour former un fin cordon cellulaire qui apparaît sombre par transparence, c’est le prolongement céphalique. La corde s’isole très tôt et permet la formation de la plaque neurale par neurulation.
Entre la 18ème et la 19ème heure, la ligne primitive atteint son maximum d’élongation (à peu près deux millimètres). L’aire pellucide ne contient plus que de l’ectoblaste en surface. D’autres cellules issues de l’ectophyle, ayant immigré dans le blastocoele secondaire, vont constituer le mésophyle puis l’endoblaste définitif refoulant l’ectoblaste. Ainsi le mésophyle va contribuer à la formation de deux ensembles distincts : le mésoderme et l’endoderme de l’embryon.
L’hypoblaste et l’entophyle donnent l’endoderme extra-embryonnaire.
A 18 heures d’incubation, tout le mésoblaste des lames latérales et extra-embryonnaires a émigré en profondeur et a divergé au niveau de la ligne primitive délaissant la région antérieure de l’aire pellucide qui reste dépourvue de mésoderme : c’est le Proamnios.
A la fin de la gastrulation, la ligne primitive représente le blastopore très allongé dont la lèvre dorsale correspond au nœud de Hensen. L’aire pellucide ne contient plus en surface que l’ectoblaste, l’épiblaste et le mésoblaste.
IV La neurulation.
La gastrulation se poursuit jusqu’à la disparition de la ligne primitive. Elle se superpose à la neurulation. A 20 heures d’incubation, apparaît un épaississement superficiel en avant du prolongement céphalique (corde). Cet allongement correspond au bord de la plaque neurale qui se soulève en constituant le repli céphalique.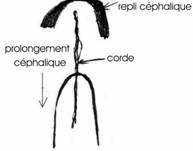
Le blastoderme s’allonge, la ligne primitive commence à régresser.
A 24 heures d’incubation, les bourrelets neuraux se soulèvent et vont à la rencontre l’un de l’autre au niveau du futur cerveau moyen.
Dans la région antérieure, on distingue une formation en doigt de gant qui est l’amorce de l’intestin antérieure. A ce stade, la région antérieure de l’aire embryonnaire s’élève au-dessus du blastoderme puis, s’accroît vers l’avant donnant naissance au repli céphalique qui coince le feuillet endodermique sous-jacent en doigt de gant. Au de là de la limite postérieure, l’intestin antérieure n’a plus de plancher et communique avec l’archantéron directement au-dessus du jaune. Le mésoderme somitique commence à se métamériser et 4 paires de somites sont nettement individualisées. A partir de la 20ème heure, une paire de somites se forme toutes les heures.
A la 26ème heure, la fusion des bourrelets neuraux débute dans la région antérieure (au niveau du cerveau). Le cerveau reste ouvert par un neuroport antérieure se fermant vers la 33ème heure d’incubation. Les bourrelets neuraux postérieurs enserrent la ligne primitive qui régresse de plus en plus. Cette région correspond au sinus rhomboïdal.
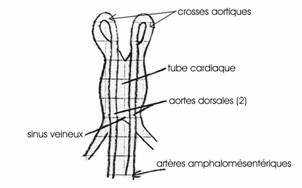
ème heure et la 33ème, le cœur va se différencier à partir d’un épaississement de la splanchnopleure. Ce tube cardiaque se poursuit vers l’arrière par 2 gros troncs veineux, les veines omphalomésentériques. Il apparaît deux aortes vers l’avant, aortes qui se replient en 2 crosses aortiques dorsales se jetant dans les 2 artères omphalomésentériques.
On a la formation d’un réseau vasculaire.
A 37 heures, le cœur bat.
A partir de la 33ème heure d’incubation, l’allure symétrique de l’embryon commence à disparaître. La partie antérieure du cerveau va se soulever et fléchir pour se tordre vers la gauche. L’embryon se soulève au-dessus du jaune puis subit une flexion dans la région antérieure et une torsion qui débute vers la 37ème heure.
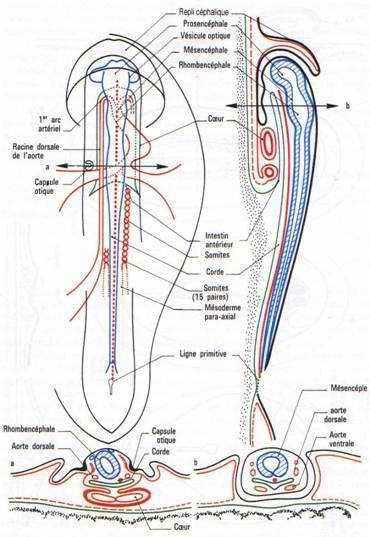
Les vésicules optiques et auditives sont nettement visibles. Un repli a pris naissance en avant de la région céphalique : c’est le capuchon amniotique qui progressera vers l’arrière, participant à la formation de l’amnios qui recouvrira l’embryon.
A ce stade, le cerveau est constitué de trois vésicules : le proencéphale, le mésencéphale et le rhombencéphale.
A 48 heures d’incubation, l’embryon qui s’est soulevé au-dessus du jaune va se coucher sur le côté gauche. La ligne primitive disparaît à 50 heures. Le capuchon amniotique recouvre la moitié de l’embryon à 50 heures. Le cerveau est formé de 5 vésicules :

Le cerveau se courbe en un V dont le sommet est occupé par le mésencéphale.
A 60 heures d’incubation, le repli amniotique recouvre les 4/5 de l’embryon. L’allantoïde apparaît sous forme d’un diverticule endodermique de l’intestin postérieur.
A 96 heures d’incubation, la cavité amniotique est complètement refermée. Le bourgeon caudal et les membres antérieurs et postérieurs sont nettement reconnaissables.
Le modelage de l’embryon va se poursuivre avec la formation des annexes embryonnaires.
V Les annexes embryonnaires.
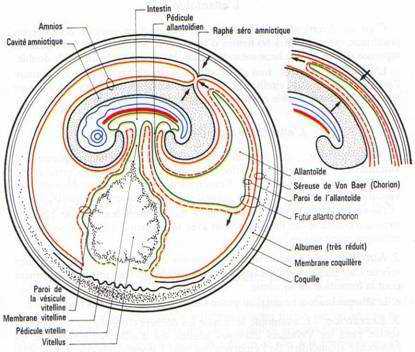
A La vésicule vitelline.
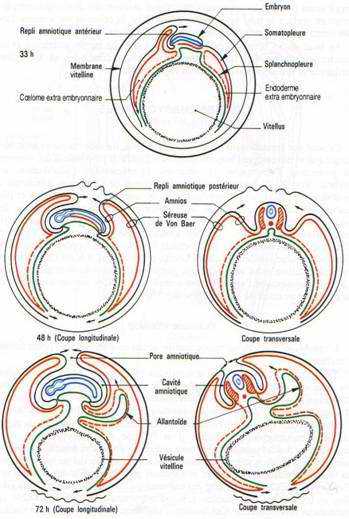
C’est le premier annexe à se former.Cette vésicule résulte de l’extension des feuillets extra-embryonnaires à la surface du jaune, constitué par de l’endoderme doublé extérieurement par le splanchnopleure. Quand l’embryon se soulève, les ébauches antérieures et postérieures du tube digestif se mettent en place. Ces ébauches iront à la rencontre l’une de l’autre au niveau de la région moyenne de l’embryon et finalement, la vésicule vitelline ne communiquera plus avec l’embryon que par un pédicule vitellin.
La vésicule vitelline, dont les parois sont fortement vascularisées, représente un organe nutritionnel extra-embryonnaire. Les cellules endodermiques, grâce à des enzymes hydrolitiques, vont transformer le vitellus en produits solubles assimilables.
B L’amnios et la séreuse.
L’amnios est une membrane qui enveloppe l’embryon comme un sac, le séparant du milieu environnant. Un repli amniotique naît dans la région antérieure puis un second va apparaître dans la région postérieure. Ils sont formés d’ectoderme doublé par de somatopleure. Quand les replis fusionnent, la partie interne des replis sera l’amnios qui limite la cavité amniotique. La partie externe, sera la séreuse. Entre la séreuse et l’amnios se trouve le coelum extra-embryonnaire qui communique avec le coelum intra-embryonnaire au niveau des lames latérales.La cavité amniotique est remplie d’une sérosité provenant de la déshydratation de l’albumen. Elle a pour rôle d’empêcher la dessiccation de l’embryon et de le protéger.
C L’allantoïde.
C’est un diverticule endodermique de l’intestin postérieur. Les invaginations de l’endoderme refoulent devant elles la splanchnopleure. L’allantoïde sera formé d’endoderme doublé extérieurement de splanchnopleure.L’allantoïde va s’entendre considérablement de manière à venir entourer tout l’amnios et la vésicule vitelline, en refoulant l’albumen vers le petit bout de l’œuf.
L’allantoïde a quatre principaux rôles :
- L’ensemble allantoïde extérieur et séreuse constitue l’allantochorion qui, extrêmement vascularisé, a fonction respiratoire (nombreux échanges gazeux).
- Le contact étroit de l’allantochorion avec la coquille permet l’absorption de sels qui seront utilisés pour l’édification du squelette de l’embryon.
- Le voisinage de l’allantoïde et du sac de l’albumen permet l’absorption du blanc : rôle nutritionnel.
- L’allantoïde stocke les produits d’excrétion du rein.
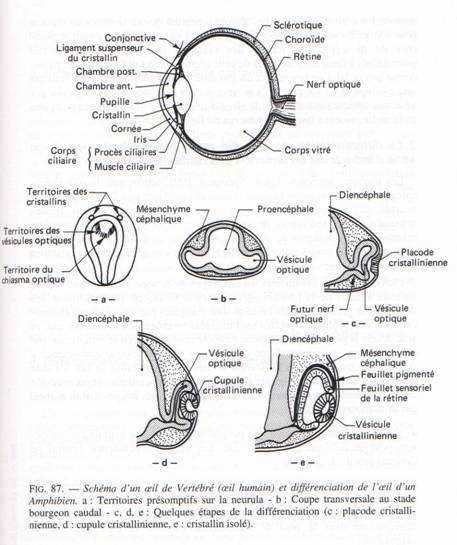
VI L’organogenèse de l’oeil.
Vers 33 heures d’incubation, les parties latérales du diencéphale s’invaginent pour donner les vésicules optiques (au nombre de 2) grâce à l’effet inducteur de la plaque précordale. Ces vésicules optiques s’allongent et se pédiculisent pour donner le nerf optique. Elles se creusent en cupules optiques formées de deux feuillets. Un mince externe à la cupule constituant la rétine pigmentaire et le feuillet interne à la cupule qui constitue la rétine nerveuse (couches de cellules sensorielles). La cupule optique va induire un épaississement de l’ectoderme. La placode cristallinienne s’invagine pour donner le cristallin formé de deux parties : un épithélium mince et vers l’intérieure et des grandes fibres cristalliniennes. Cela permet la transformation de l’ectoderme en cornée transparente.